|
|
|
Goldstein Lab
Research (since ~2006)
Overview
When asked whether I am a theorist or
an experimentalist, my reply is that I am a scientist.
Our group seeks to understand fundamental principles that govern the
behavior of nonequilibrium
systems in physics and biology, using a combination of experiment and
theory.
This research is not easily described by a single, conventional
academic label; rather, it involves
the domains of condensed matter physics, physical chemistry, biological
physics, fluid dynamics,
applied mathematics, and geophysics. I subscribe to
Poincaré's motivation:
The scientist does not study nature because it is useful;
he studies it because he delights in
it, and he delights in it because
it is beautiful. If nature were not
beautiful, it would not be worth knowing, and
if nature were not worth knowing,
life would not be worth living.
I also believe that some of the best science is close to art, and that
Glenn Gould captured this spirit when he said
The purpose of art is not the release of a momentary ejection of
adrenaline but rather the gradual,
lifelong construction of a state of wonder and serenity.
Current research in my group falls into two broad categories:
Biological Physics and
Natural Pattern Formation.
I. Biological Physics
Our group is currently focused on a range of questions centred around the
origins of multicellularity. We use extensively the Volvocine green algae as a
class of model organisms to understand the driving force behind the emergence of
germ-soma differentiation, the nature of flagellar synchronization, the mechanisms
of phototaxis, and basic aspects of biological fluid dynamics. A parallel set of
investigations on the phenomenon of cytoplasmic streaming aims to answer the basic
question of its biological purpose. Here we use the Characean algae as model organisms,
and also have a developing collaboration involving streaming in the developing fruit fly
oocyte.
Swimming, Stirring, and Scaling in the Volvocales
 One of the most fundamental issues in biology is the nature of evolutionary transitions
from single cell organisms to multicellular ones. It is a general rule of nature that larger organisms
are more complex, at least as measured by the number of distinct types of cells present.
This reflects the fitness advantage conferred by a division of labor among specialized cells
over homogeneous totipotency. Yet, increasing size has both costs and benefits, and the search for
the driving forces behind the evolution of multicellularity is becoming a very
active area of research.
Not surprisingly for microscopic life in
a fluid environment, many of the processes involved are related to transport and locomotion,
for efficient exchange of chemical species with the environment is one of the most basic
features of life. We
study these issues using as model organisms the Volvocales, which comprise algae
ranging from Chlamydomonas (A),
swimming single cells, to coenobia of undifferentiated cells, such as Gonium (B) and
Pandorina (C), to Volvox sp. (E,F), where the surface is
covered by thousands of flagellated somatic cells, while the interior
contains a far smaller number of gonidia. The existence of these closely related species
allows one to study some of the most basic questions in the evolution of multicellularity.
What determines the length scale or the cell number at which germ-soma differentiation appears?
How do simple multicellular (or colonial) organisms lacking a central nervous system achieve
the apparently coordinated motion they exhibit? What developmental program leads to the
long-range cellular orientation observed in the larger species?
Our first two works in this area laid the groundwork for many of these issues by developing
micromanipulation and fluid dynamical methods of studying the flagella-driven flows in
these organisms, thereby establishing
the high Péclet numbers found in these systems and showing
through experiment and theory that such flows can remove what would otherwise be a diffusional
bottleneck in nutrient uptake limiting viability of the larger species.
One of the most fundamental issues in biology is the nature of evolutionary transitions
from single cell organisms to multicellular ones. It is a general rule of nature that larger organisms
are more complex, at least as measured by the number of distinct types of cells present.
This reflects the fitness advantage conferred by a division of labor among specialized cells
over homogeneous totipotency. Yet, increasing size has both costs and benefits, and the search for
the driving forces behind the evolution of multicellularity is becoming a very
active area of research.
Not surprisingly for microscopic life in
a fluid environment, many of the processes involved are related to transport and locomotion,
for efficient exchange of chemical species with the environment is one of the most basic
features of life. We
study these issues using as model organisms the Volvocales, which comprise algae
ranging from Chlamydomonas (A),
swimming single cells, to coenobia of undifferentiated cells, such as Gonium (B) and
Pandorina (C), to Volvox sp. (E,F), where the surface is
covered by thousands of flagellated somatic cells, while the interior
contains a far smaller number of gonidia. The existence of these closely related species
allows one to study some of the most basic questions in the evolution of multicellularity.
What determines the length scale or the cell number at which germ-soma differentiation appears?
How do simple multicellular (or colonial) organisms lacking a central nervous system achieve
the apparently coordinated motion they exhibit? What developmental program leads to the
long-range cellular orientation observed in the larger species?
Our first two works in this area laid the groundwork for many of these issues by developing
micromanipulation and fluid dynamical methods of studying the flagella-driven flows in
these organisms, thereby establishing
the high Péclet numbers found in these systems and showing
through experiment and theory that such flows can remove what would otherwise be a diffusional
bottleneck in nutrient uptake limiting viability of the larger species.
[back to overview]
Tracking Protists in Three Dimensions
 Many flagellated protists display
swimming behavior that is inherently three dimensional, and a number of important questions
in biology and physics are associated with how the motion of such organisms is
related to their body plan and to external stimuli such as
light, dissolved molecular species, gravity, temperature,
boundaries, and electromagnetic fields. It is thus desirable to
track their position and orientation in 3D with high spatiotemporal
resolution and, unless desired, free from systematic
bias introduced by external stimuli, background fluid
motion, and hydrodynamic surface effects.
We have developed such an apparatus, optimized for
tracking swimming micro-organisms in the size range of
10-1000 microns, in three dimensions, far from surfaces, and with negligible background
convective fluid motion. Charge coupled device cameras attached to two long working distance
microscopes synchronously image the sample from two perpendicular directions, with narrow band
dark-field or bright-field illumination chosen to avoid triggering a phototactic response. The images
from the two cameras can be combined to yield 3D tracks of the organism. Using additional, highly
directional broad-spectrum illumination with millisecond timing control the phototactic trajectories
in 3D of organisms ranging from
Chlamydomonas to
Volvox
can be studied in detail.
Surface-mediated hydrodynamic interactions can also be investigated without convective
interference. Minimal modifications to the apparatus allow for studies of chemotaxis and other
taxes.
Many flagellated protists display
swimming behavior that is inherently three dimensional, and a number of important questions
in biology and physics are associated with how the motion of such organisms is
related to their body plan and to external stimuli such as
light, dissolved molecular species, gravity, temperature,
boundaries, and electromagnetic fields. It is thus desirable to
track their position and orientation in 3D with high spatiotemporal
resolution and, unless desired, free from systematic
bias introduced by external stimuli, background fluid
motion, and hydrodynamic surface effects.
We have developed such an apparatus, optimized for
tracking swimming micro-organisms in the size range of
10-1000 microns, in three dimensions, far from surfaces, and with negligible background
convective fluid motion. Charge coupled device cameras attached to two long working distance
microscopes synchronously image the sample from two perpendicular directions, with narrow band
dark-field or bright-field illumination chosen to avoid triggering a phototactic response. The images
from the two cameras can be combined to yield 3D tracks of the organism. Using additional, highly
directional broad-spectrum illumination with millisecond timing control the phototactic trajectories
in 3D of organisms ranging from
Chlamydomonas to
Volvox
can be studied in detail.
Surface-mediated hydrodynamic interactions can also be investigated without convective
interference. Minimal modifications to the apparatus allow for studies of chemotaxis and other
taxes.
[back to overview]
Hydrodynamic Bound States of Swimming Algae
 Because of its bottom-heaviness, in the absence of phototactic cues
Volvox swims
upward against gravity. Using glass chambers we
discovered that when nearby
colonies reach the chamber ceiling they are attracted together and can form a stable bound
state in which they "waltz" around each other. The attractive interaction was shown to
be a surface-mediated effect associated with the density offset of the colonies relative
to water (so that in the far-field each is described by a downward-pointing Stokeslet), in the
presence of a no-slip wall. Quantitative agreement with experimental observations on infalling
trajectories was achieved with no free parameters.
Lubrication theory for the dynamics of nearby spinning, bottom-heavy
colonies can be used to explain the orbiting dynamics of the bound states. A second "minueting"
dynamics occurs with older colonies that hover near the lower chamber wall.
These phenomena are suggested to
underlie observed clustering of Volvox at surfaces.
Because of its bottom-heaviness, in the absence of phototactic cues
Volvox swims
upward against gravity. Using glass chambers we
discovered that when nearby
colonies reach the chamber ceiling they are attracted together and can form a stable bound
state in which they "waltz" around each other. The attractive interaction was shown to
be a surface-mediated effect associated with the density offset of the colonies relative
to water (so that in the far-field each is described by a downward-pointing Stokeslet), in the
presence of a no-slip wall. Quantitative agreement with experimental observations on infalling
trajectories was achieved with no free parameters.
Lubrication theory for the dynamics of nearby spinning, bottom-heavy
colonies can be used to explain the orbiting dynamics of the bound states. A second "minueting"
dynamics occurs with older colonies that hover near the lower chamber wall.
These phenomena are suggested to
underlie observed clustering of Volvox at surfaces.
[back to overview]
Synchronization of Eukaryotic Flagella
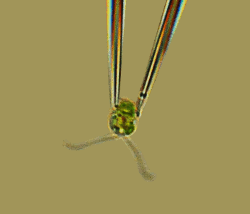 The coordination of eukaryotic flagella is essential for many of the most basic processes of life
(motility, sensing, and development), yet its emergence and regulation and its connection to
locomotion are poorly understood. Previous studies show that the unicellular alga
and Chlamydomonas reinhardtii,
widely regarded as an ideal system in which to study flagellar biology, swims forward by the
synchronous action of its two flagella. Using high-speed imaging over long intervals, we
found a
richer behavior: A cell swimming in the dark stochastically switches between synchronous and
asynchronous flagellar beating.
The synchronous state is interrupted stochastically
by phase slips. The dynamics of slips and the statistics of phase-locked intervals are
consistent
with a low-dimensional
stochastic model of hydrodynamically coupled oscillators, with a noise amplitude set by the
intrinsic fluctuations of single flagellar beats.
Three-dimensional tracking shows that the alternation between synchronous and asynchronous
beating regimes leads,
respectively, to nearly straight swimming and to abrupt large reorientations, which yield a
eukaryotic version of the "run-and-tumble" motion of peritrichously flagellated bacteria.
The coordination of eukaryotic flagella is essential for many of the most basic processes of life
(motility, sensing, and development), yet its emergence and regulation and its connection to
locomotion are poorly understood. Previous studies show that the unicellular alga
and Chlamydomonas reinhardtii,
widely regarded as an ideal system in which to study flagellar biology, swims forward by the
synchronous action of its two flagella. Using high-speed imaging over long intervals, we
found a
richer behavior: A cell swimming in the dark stochastically switches between synchronous and
asynchronous flagellar beating.
The synchronous state is interrupted stochastically
by phase slips. The dynamics of slips and the statistics of phase-locked intervals are
consistent
with a low-dimensional
stochastic model of hydrodynamically coupled oscillators, with a noise amplitude set by the
intrinsic fluctuations of single flagellar beats.
Three-dimensional tracking shows that the alternation between synchronous and asynchronous
beating regimes leads,
respectively, to nearly straight swimming and to abrupt large reorientations, which yield a
eukaryotic version of the "run-and-tumble" motion of peritrichously flagellated bacteria.
[back to overview]
Fidelity of Adaptive Phototaxis
 Along the evolutionary path from single cells to multicellular organisms
with a central nervous system are species of intermediate
complexity that move in ways suggesting high-level coordination,
yet have none. Instead, organisms of this type possess many autonomous
cells endowed with programs that have evolved to achieve
concerted responses to environmental stimuli. In our recent
paper experiment
and theory are used to develop a quantitative understanding of
how cells of such organisms coordinate to achieve phototaxis,
by using the colonial alga Volvox carteri as a model. It is shown
that the surface somatic cells act as individuals but are orchestrated
by their relative position in the spherical extracellular matrix and
their common photoresponse function to achieve colony-level coordination.
Analysis of models that range from the minimal to the
biologically faithful shows that, because the flagellar beating displays
an adaptive down-regulation in response to light, the colony
needs to spin around its swimming direction and that the response
kinetics and natural spinning frequency of the colony appear to be
mutually tuned to give the maximum photoresponse. These
models further predict that the phototactic ability decreases dramatically
when the colony does not spin at its natural frequency,
a result confirmed by phototaxis assays in which colony rotation
was slowed by increasing the fluid viscosity.
Along the evolutionary path from single cells to multicellular organisms
with a central nervous system are species of intermediate
complexity that move in ways suggesting high-level coordination,
yet have none. Instead, organisms of this type possess many autonomous
cells endowed with programs that have evolved to achieve
concerted responses to environmental stimuli. In our recent
paper experiment
and theory are used to develop a quantitative understanding of
how cells of such organisms coordinate to achieve phototaxis,
by using the colonial alga Volvox carteri as a model. It is shown
that the surface somatic cells act as individuals but are orchestrated
by their relative position in the spherical extracellular matrix and
their common photoresponse function to achieve colony-level coordination.
Analysis of models that range from the minimal to the
biologically faithful shows that, because the flagellar beating displays
an adaptive down-regulation in response to light, the colony
needs to spin around its swimming direction and that the response
kinetics and natural spinning frequency of the colony appear to be
mutually tuned to give the maximum photoresponse. These
models further predict that the phototactic ability decreases dramatically
when the colony does not spin at its natural frequency,
a result confirmed by phototaxis assays in which colony rotation
was slowed by increasing the fluid viscosity.
[back to overview]
Emergence of Synchronized Beating During the Regrowth of Eukaryotic Flagella
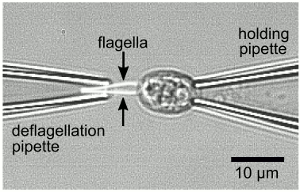 A fundamental issue in the biology of eukaryotic flagella is the origin of synchronized beating
observed in tissues and organisms containing multiple flagella. Our recent studies of the
biflagellate unicellular alga Chlamydomonas reinhardtii provided the first evidence that
the interflagellar coupling responsible for synchronization is of hydrodynamic origin. To
investigate this mechanism in detail we have
studied
synchronization in Chlamydomonas as
its flagella slowly regrow after mechanically-induced self-scission. The duration of synchronized
intervals is found to be strongly dependent on flagellar length. Analysis within a stochastic model
of coupled phase oscillators is used to extract the length dependence of the interflagellar coupling
and the intrinsic beat frequencies of the two flagella. Physical and biological considerations that
may explain these results are proposed.
A fundamental issue in the biology of eukaryotic flagella is the origin of synchronized beating
observed in tissues and organisms containing multiple flagella. Our recent studies of the
biflagellate unicellular alga Chlamydomonas reinhardtii provided the first evidence that
the interflagellar coupling responsible for synchronization is of hydrodynamic origin. To
investigate this mechanism in detail we have
studied
synchronization in Chlamydomonas as
its flagella slowly regrow after mechanically-induced self-scission. The duration of synchronized
intervals is found to be strongly dependent on flagellar length. Analysis within a stochastic model
of coupled phase oscillators is used to extract the length dependence of the interflagellar coupling
and the intrinsic beat frequencies of the two flagella. Physical and biological considerations that
may explain these results are proposed.
Anomalous Tracer Statistics in Active Suspensions
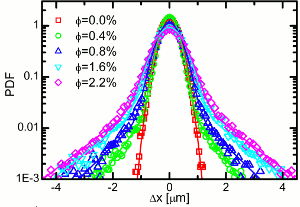 In contexts such as suspension feeding in marine ecologies there is an interplay between Brownian
motion of nonmotile particles and their advection by flows from swimming microorganisms.
One appealing point of view is that the sea of swimming organisms constitutes an effective "thermal
bath" analogous to the multitudes of molecules responsible
for Brownian motion, where each encounter of a
tracer particle with a swimmer provides a random kick.
In conventional Brownian motion, e.g., with micron-size
particles in water, there is an enormous separation of time
scales between the duration of molecular collisions (ps)
and the observed particle motion (ms). In contrast, in a
suspension of microorganisms it is possible to resolve the
encounters with tracer particles, and the dynamical problem
involves correlated advective trajectories in the presence
of true Brownian noise.
As a laboratory realization of this, we have
studied
passive tracers in suspensions of eukaryotic swimmers, the alga
Chlamydomonas reinhardtii.
While the cells behave ballistically over short intervals, the tracers behave
diffusively, with a time-dependent but self-similar probability distribution function of displacements
consisting of a Gaussian core and robust exponential tails. We emphasize the role of flagellar beating in
creating oscillatory flows that exceed Brownian motion far from each swimmer.
In contexts such as suspension feeding in marine ecologies there is an interplay between Brownian
motion of nonmotile particles and their advection by flows from swimming microorganisms.
One appealing point of view is that the sea of swimming organisms constitutes an effective "thermal
bath" analogous to the multitudes of molecules responsible
for Brownian motion, where each encounter of a
tracer particle with a swimmer provides a random kick.
In conventional Brownian motion, e.g., with micron-size
particles in water, there is an enormous separation of time
scales between the duration of molecular collisions (ps)
and the observed particle motion (ms). In contrast, in a
suspension of microorganisms it is possible to resolve the
encounters with tracer particles, and the dynamical problem
involves correlated advective trajectories in the presence
of true Brownian noise.
As a laboratory realization of this, we have
studied
passive tracers in suspensions of eukaryotic swimmers, the alga
Chlamydomonas reinhardtii.
While the cells behave ballistically over short intervals, the tracers behave
diffusively, with a time-dependent but self-similar probability distribution function of displacements
consisting of a Gaussian core and robust exponential tails. We emphasize the role of flagellar beating in
creating oscillatory flows that exceed Brownian motion far from each swimmer.
[back to overview]
Direct Measurements of the Flow Fields Around Swimming Microorganisms
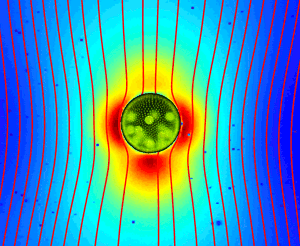 Swimming microorganisms create flows that influence their mutual interactions and modify the
rheology of their suspensions. While extensively studied theoretically, these flows have not been measured
in detail around any freely-swimming microorganism. Using a synthesis of tracking microscopy,
particle imaging velocimetry, and particle tracking velocimetry, we
achieved
such measurements
for the microphytes
Volvox carteri,
and Chlamydomonas reinhardtii.
The minute (~0.3%) density excess of
V. carteri
over water leads to a strongly dominant Stokeslet contribution, with the widely-assumed stresslet flow only a
correction to the subleading source dipole term. This implies that suspensions of
V. carteri
have features
similar to suspensions of sedimenting particles (see the next section for an explanation of
the consequences of this). The flow in the region around
C. reinhardtii
where
significant hydrodynamic interaction is likely to occur differs qualitatively from a puller stresslet, and can
be described by a simple three-Stokeslet model.
Swimming microorganisms create flows that influence their mutual interactions and modify the
rheology of their suspensions. While extensively studied theoretically, these flows have not been measured
in detail around any freely-swimming microorganism. Using a synthesis of tracking microscopy,
particle imaging velocimetry, and particle tracking velocimetry, we
achieved
such measurements
for the microphytes
Volvox carteri,
and Chlamydomonas reinhardtii.
The minute (~0.3%) density excess of
V. carteri
over water leads to a strongly dominant Stokeslet contribution, with the widely-assumed stresslet flow only a
correction to the subleading source dipole term. This implies that suspensions of
V. carteri
have features
similar to suspensions of sedimenting particles (see the next section for an explanation of
the consequences of this). The flow in the region around
C. reinhardtii
where
significant hydrodynamic interaction is likely to occur differs qualitatively from a puller stresslet, and can
be described by a simple three-Stokeslet model.
[back to overview]
Fluid Dynamics and Noise in Bacterial Cell-Cell and Cell-Surface Interactions
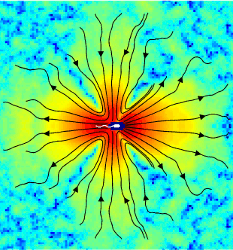 Bacterial processes ranging from gene expression to motility and
biofilm formation are constantly challenged by internal and external
noise. While the importance of stochastic fluctuations has been
appreciated for chemotaxis, it is currently believed that deterministic
long-range fluid dynamical effects govern cell-cell and cell-surface scattering,
the elementary events that lead to swarming
and collective swimming in active suspensions and to the formation
of biofilms. We have succeeded in making the first direct
measurements
of the
bacterial flow field generated by individual swimming Escherichia
coli both far from and near to a solid surface. These experiments
allowed us to examine the relative importance of fluid dynamics
and rotational diffusion for bacteria. For cell-cell interactions it
is shown that thermal and intrinsic stochasticity drown the effects
of long-range fluid dynamics, implying that physical interactions
between bacteria are determined by steric collisions and near-field
lubrication forces. This dominance of short-range forces closely
links collective motion in bacterial suspensions to self-organization
in driven granular systems, assemblages of biofilaments, and animal
flocks. For the scattering of bacteria with surfaces, long-range
fluid dynamical interactions are also shown to be negligible before
collisions; however, once the bacterium swims along the surface
within a few microns after an aligning collision, hydrodynamic
effects can contribute to the experimentally observed, long residence
times. Because these results are based on purely mechanical
properties, they apply to a wide range of microorganisms.
Bacterial processes ranging from gene expression to motility and
biofilm formation are constantly challenged by internal and external
noise. While the importance of stochastic fluctuations has been
appreciated for chemotaxis, it is currently believed that deterministic
long-range fluid dynamical effects govern cell-cell and cell-surface scattering,
the elementary events that lead to swarming
and collective swimming in active suspensions and to the formation
of biofilms. We have succeeded in making the first direct
measurements
of the
bacterial flow field generated by individual swimming Escherichia
coli both far from and near to a solid surface. These experiments
allowed us to examine the relative importance of fluid dynamics
and rotational diffusion for bacteria. For cell-cell interactions it
is shown that thermal and intrinsic stochasticity drown the effects
of long-range fluid dynamics, implying that physical interactions
between bacteria are determined by steric collisions and near-field
lubrication forces. This dominance of short-range forces closely
links collective motion in bacterial suspensions to self-organization
in driven granular systems, assemblages of biofilaments, and animal
flocks. For the scattering of bacteria with surfaces, long-range
fluid dynamical interactions are also shown to be negligible before
collisions; however, once the bacterium swims along the surface
within a few microns after an aligning collision, hydrodynamic
effects can contribute to the experimentally observed, long residence
times. Because these results are based on purely mechanical
properties, they apply to a wide range of microorganisms.
[back to overview]
Fluid Velocity Fluctuations in a Suspension of Swimming Protists
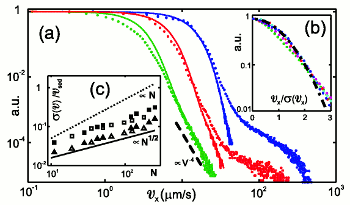 In dilute suspensions of swimming microorganisms the local fluid velocity is a
random superposition of the flow fields set up by the individual organisms, which in turn
have multipole contributions decaying as inverse powers of distance from the organism.
We have studied the relationship between the decay exponent of the dominant multipole contribution and
the statistics of velocity fluctuations, and
derived
the conditions under which the central limit theorem guarantees a Gaussian probability distribution function
of velocities are satisied. This holds when the leading force singularity is a Stokeslet, but
not when it is any higher multipole. These results are confirmed by numerical studies and by
experiments on suspensions of the alga
Volvox carteri,
which show that deviations from Gaussianity arise from near-field effects. Such observations
are complementary to the observation (see above) of non-Gaussian finite-time particle displacement
distribution functions in suspensions of
C. reinhardtii.
In dilute suspensions of swimming microorganisms the local fluid velocity is a
random superposition of the flow fields set up by the individual organisms, which in turn
have multipole contributions decaying as inverse powers of distance from the organism.
We have studied the relationship between the decay exponent of the dominant multipole contribution and
the statistics of velocity fluctuations, and
derived
the conditions under which the central limit theorem guarantees a Gaussian probability distribution function
of velocities are satisied. This holds when the leading force singularity is a Stokeslet, but
not when it is any higher multipole. These results are confirmed by numerical studies and by
experiments on suspensions of the alga
Volvox carteri,
which show that deviations from Gaussianity arise from near-field effects. Such observations
are complementary to the observation (see above) of non-Gaussian finite-time particle displacement
distribution functions in suspensions of
C. reinhardtii.
[back to overview]
Physics and Biology of Cytoplasmic Streaming
 Since Bonaventura Corti's discovery in 1774 of the persistent
circulation of the cytoplasm of plant cells, the phenomenon
now known as cytoplasmic streaming or cyclosis has
been conjectured to play an important role in metabolism. It
occurs in organisms as diverse as amoebae, algae and
terrestrial plants, and fungi. In plants it is driven
by multitudes of the motor protein myosin moving along bundled
actin at the boundary of the cytoplasm, carrying microscopic
particles or organelles, and entraining fluid. The motion
of protoplasmic granules entrained in the flow includes unidirectional
streaming, "fountain streaming" (in which the motion
near the central axis of the cell is opposite to that near the
periphery), and spiral "rotational streaming." The fact
that transport by fluid motion becomes necessary to outrun the
slow pace of diffusion in larger organisms, as emphasized in the
celebrated essay by Haldane on size in biology, has been a
theme in discussions of cytoplasmic streaming for many years.
Yet, there has been little theoretical work and fewer experiments
that have quantified the full implications of cytoplasmic streaming
for transport and mixing. We have embarked on a research program aimed
at answering some of the most basic open questions in this field:
What purpose does cytoplasmic streaming have in cells? How does it impact on homeostasis
and development? What gives rise to the often complex flow geometries found
in streaming?
Since Bonaventura Corti's discovery in 1774 of the persistent
circulation of the cytoplasm of plant cells, the phenomenon
now known as cytoplasmic streaming or cyclosis has
been conjectured to play an important role in metabolism. It
occurs in organisms as diverse as amoebae, algae and
terrestrial plants, and fungi. In plants it is driven
by multitudes of the motor protein myosin moving along bundled
actin at the boundary of the cytoplasm, carrying microscopic
particles or organelles, and entraining fluid. The motion
of protoplasmic granules entrained in the flow includes unidirectional
streaming, "fountain streaming" (in which the motion
near the central axis of the cell is opposite to that near the
periphery), and spiral "rotational streaming." The fact
that transport by fluid motion becomes necessary to outrun the
slow pace of diffusion in larger organisms, as emphasized in the
celebrated essay by Haldane on size in biology, has been a
theme in discussions of cytoplasmic streaming for many years.
Yet, there has been little theoretical work and fewer experiments
that have quantified the full implications of cytoplasmic streaming
for transport and mixing. We have embarked on a research program aimed
at answering some of the most basic open questions in this field:
What purpose does cytoplasmic streaming have in cells? How does it impact on homeostasis
and development? What gives rise to the often complex flow geometries found
in streaming?
[back to overview]
Microfluidics of Cytoplasmic Streaming
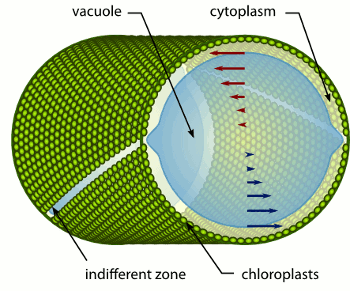 In the more than two centuries since its discovery, streaming
has frequently been conjectured to aid in transport and mixing of
molecular species in the cytoplasm and, by implication, in cellular
homeostasis, yet no theoretical analysis has been presented to quantify
these processes. We show by solution of the coupled dynamics
of fluid flow and diffusion appropriate to the archetypal "rotational
streaming" of algal species such as Chara and Nitella that internal
mixing and the transient dynamical response to changing external
conditions can indeed be enhanced by streaming, but to an extent
that depends strongly on the pitch of the helical flow. The possibility
that this may have a developmental consequence is illustrated by the
coincidence of the exponential growth phase of Nitella and the point
of maximum enhancement of those processes.
Key to these results is the discovery and analysis
of a circulatory flow transverse
to the cylinder's long axis, akin to Dean vortices at finite Reynolds numbers, which arises from the chiral
geometry. Strongly enhanced lateral transport and longitudinal homogenization occur if the transverse
Péclet number is sufficiently large, with scaling laws arising from boundary layers.
In the more than two centuries since its discovery, streaming
has frequently been conjectured to aid in transport and mixing of
molecular species in the cytoplasm and, by implication, in cellular
homeostasis, yet no theoretical analysis has been presented to quantify
these processes. We show by solution of the coupled dynamics
of fluid flow and diffusion appropriate to the archetypal "rotational
streaming" of algal species such as Chara and Nitella that internal
mixing and the transient dynamical response to changing external
conditions can indeed be enhanced by streaming, but to an extent
that depends strongly on the pitch of the helical flow. The possibility
that this may have a developmental consequence is illustrated by the
coincidence of the exponential growth phase of Nitella and the point
of maximum enhancement of those processes.
Key to these results is the discovery and analysis
of a circulatory flow transverse
to the cylinder's long axis, akin to Dean vortices at finite Reynolds numbers, which arises from the chiral
geometry. Strongly enhanced lateral transport and longitudinal homogenization occur if the transverse
Péclet number is sufficiently large, with scaling laws arising from boundary layers.
[back to overview]
Measurement of Cytoplasmic Streaming by Magnetic Resonance Velocimetry
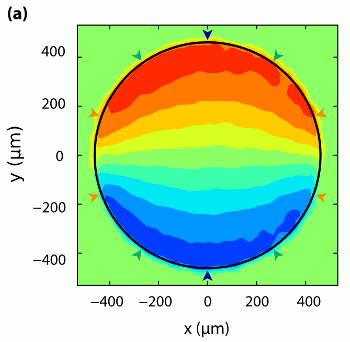 In the giant cylindrical cells found in Characean algae, multitudes of the molecular
motor myosin transport the cytoplasm along opposing spiralling bands covering the
inside of the cell wall, generating a helical shear flow in the large central vacuole.
It has been suggested that such flows enhance mixing
within the vacuole and
thereby play a role in regulating metabolism. For this to occur the membrane that
encloses the vacuole, namely the tonoplast, must transmit efficiently the hydrodynamic
shear generated in the cytoplasm. Existing measurements of streaming flows are of
insufficient spatial resolution and extent to provide tests of fluid mechanical theories
of such flows and information on the shear transmission. We have used magnetic
resonance velocimetry (MRV) to obtain the first measurements of cytoplasmic
streaming velocities in single living cells. The spatial variation of the longitudinal
velocity field in cross-sections of internodal cells of Chara corallina was obtained
and shown to be in quantitative agreement with our theoretical
analysis of rotational cytoplasmic streaming driven by bidirectional
helical forcing in the cytoplasm, with direct shear transmission by the tonoplast.
In the giant cylindrical cells found in Characean algae, multitudes of the molecular
motor myosin transport the cytoplasm along opposing spiralling bands covering the
inside of the cell wall, generating a helical shear flow in the large central vacuole.
It has been suggested that such flows enhance mixing
within the vacuole and
thereby play a role in regulating metabolism. For this to occur the membrane that
encloses the vacuole, namely the tonoplast, must transmit efficiently the hydrodynamic
shear generated in the cytoplasm. Existing measurements of streaming flows are of
insufficient spatial resolution and extent to provide tests of fluid mechanical theories
of such flows and information on the shear transmission. We have used magnetic
resonance velocimetry (MRV) to obtain the first measurements of cytoplasmic
streaming velocities in single living cells. The spatial variation of the longitudinal
velocity field in cross-sections of internodal cells of Chara corallina was obtained
and shown to be in quantitative agreement with our theoretical
analysis of rotational cytoplasmic streaming driven by bidirectional
helical forcing in the cytoplasm, with direct shear transmission by the tonoplast.
[back to overview]
Shear-Driven Circulation Patterns in Lipid Membrane Vesicles
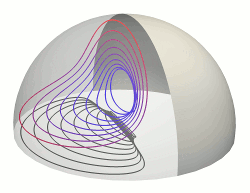 Recent experiments have shown that when a hemispherical lipid vesicle attached to a
solid surface is subjected to a simple shear flow it exhibits a pattern of membrane circulation
much like a dipole vortex. This is in marked contrast to the toroidal circulation
that would occur in the related problem of a drop of immiscible fluid attached to a surface
and subjected to shear. This profound difference in floow patterns arises from the
lateral incompressibility of the membrane, which restricts the observable flows to those
in which the velocity field in the membrane is two-dimensionally divergence free, so there is
no return flow to the bulk. We have studied
these circulation patterns within the simplest model of membrane
fluid dynamics. A systematic expansion of the flow field based on Papkovich-Neuber potentials is
developed for general viscosity ratios between the membrane and the surrounding fluids.
Comparison with experimental results [C. Vezy, G. Massiera, and A. Viallat, Soft Matter
3, 844 (2007)] is made, and it is shown how such studies can allow measurements of the
membrane viscosity. Issues of symmetry-breaking and pattern selection are discussed.
Recent experiments have shown that when a hemispherical lipid vesicle attached to a
solid surface is subjected to a simple shear flow it exhibits a pattern of membrane circulation
much like a dipole vortex. This is in marked contrast to the toroidal circulation
that would occur in the related problem of a drop of immiscible fluid attached to a surface
and subjected to shear. This profound difference in floow patterns arises from the
lateral incompressibility of the membrane, which restricts the observable flows to those
in which the velocity field in the membrane is two-dimensionally divergence free, so there is
no return flow to the bulk. We have studied
these circulation patterns within the simplest model of membrane
fluid dynamics. A systematic expansion of the flow field based on Papkovich-Neuber potentials is
developed for general viscosity ratios between the membrane and the surrounding fluids.
Comparison with experimental results [C. Vezy, G. Massiera, and A. Viallat, Soft Matter
3, 844 (2007)] is made, and it is shown how such studies can allow measurements of the
membrane viscosity. Issues of symmetry-breaking and pattern selection are discussed.
[back to overview]
Fluctuations, Dynamics, and the Stretch-Coil Transition of
Single Actin Filaments in Extensional Flows
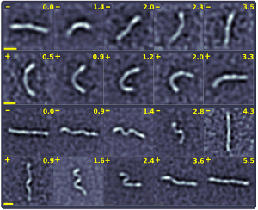 Recent work on the motion of elastic filaments subject
to hydrodynamic forces has revealed complex nonlinear
dynamics in the neighborhood of hyperbolic stagnation
points in the flow. Unlike the simpler orbits of rigid
elongated objects in the presence of shear and vorticity,
these dynamics arise from the tension induced in the filament by an extensional
flow, which beyond a critical
value can induce an instability analogous to Euler buckling of a filament with thrust at its
two ends. This predicted `stretch-coil' transition, which is complementary to the
`coil-stretch' transition of
exible polymers, has recently been observed with macroscopic fibers
in cellular flows generated by electrodynamic forcing.
Motivated by the role that semiflexible polymers subject to hydrodynamic forcing play in cytoskeletal
motions in the cell, particularly when filaments guide molecular motors whose motions create
flows, we have used a microfluidic cross-flow geometry
to provide the first comprehensive study of the interplay between
tension, fluctuations, and buckling of biopolymers, including a
fluctuation-rounded stretch-coil transition of actin filaments.
Recent work on the motion of elastic filaments subject
to hydrodynamic forces has revealed complex nonlinear
dynamics in the neighborhood of hyperbolic stagnation
points in the flow. Unlike the simpler orbits of rigid
elongated objects in the presence of shear and vorticity,
these dynamics arise from the tension induced in the filament by an extensional
flow, which beyond a critical
value can induce an instability analogous to Euler buckling of a filament with thrust at its
two ends. This predicted `stretch-coil' transition, which is complementary to the
`coil-stretch' transition of
exible polymers, has recently been observed with macroscopic fibers
in cellular flows generated by electrodynamic forcing.
Motivated by the role that semiflexible polymers subject to hydrodynamic forcing play in cytoskeletal
motions in the cell, particularly when filaments guide molecular motors whose motions create
flows, we have used a microfluidic cross-flow geometry
to provide the first comprehensive study of the interplay between
tension, fluctuations, and buckling of biopolymers, including a
fluctuation-rounded stretch-coil transition of actin filaments.
[back to overview]
Coupling of Active Motion and Advection Shapes Intracellular Cargo Transport
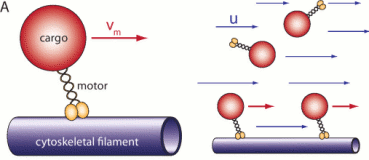 Intracellular cargo transport can arise from passive diffusion, active motor-driven transport
along cytoskeletal filament networks, and passive advection by fluid flows entrained by such
motor/cargo motion. Active and advective transport are thus intrinsically coupled as related,
yet different representations of the same underlying network structure. We have used
a reaction-advection-diffusion system to show that this
coupling affects the transport and
localization of a passive tracer in a confined geometry. For sufficiently low diffusion,
cargo localization to a target zone is optimized either by low reaction kinetics and decoupling of
bound and unbound states, or by a mostly disordered cytoskeletal network with only weak
directional bias. These generic results may help to rationalize subtle features of cytoskeletal networks, for example as observed for microtubules in fly oocytes.
Intracellular cargo transport can arise from passive diffusion, active motor-driven transport
along cytoskeletal filament networks, and passive advection by fluid flows entrained by such
motor/cargo motion. Active and advective transport are thus intrinsically coupled as related,
yet different representations of the same underlying network structure. We have used
a reaction-advection-diffusion system to show that this
coupling affects the transport and
localization of a passive tracer in a confined geometry. For sufficiently low diffusion,
cargo localization to a target zone is optimized either by low reaction kinetics and decoupling of
bound and unbound states, or by a mostly disordered cytoskeletal network with only weak
directional bias. These generic results may help to rationalize subtle features of cytoskeletal networks, for example as observed for microtubules in fly oocytes.
[back to overview]
Cytoplasmic Streaming in Drosophila Oocytes Varies with Kinesin
Activity and Correlates with the Microtubule Cytoskeleton Architecture
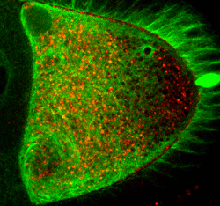 Cells can localize molecules asymmetrically through the combined action of cytoplasmic streaming,
which circulates their fluid contents, and specific anchoring mechanisms.
Streaming also contributes to the distribution of nutrients and organelles such as
chloroplasts in plants, the asymmetric position of the meiotic spindle in mammalian embryos,
and the developmental potential of the zygote, yet little has been known quantitatively about the relationship
between streaming and the motor activity which drives it.
In a collaboration with Dr. Isabel Palacios and her student Dr. Lucy S. Williams (Zoology, Cambridge)
we have used Particle Image Velocimetry (PIV)
to quantify the statistical properties of Kinesin-dependent
streaming during mid-oogenesis in Drosophila.
We find that streaming can be used to detect subtle changes in Kinesin activity and
that the flows reflect the architecture of the microtubule cytoskeleton.
Furthermore, based on characterization of the rheology of the cytoplasm in vivo, we
establish estimates of the number of Kinesins required to drive the observed streaming.
Using this in vivo data as the basis of a model for transport, we suggest that the disordered
character of transport at mid-oogenesis, as revealed by streaming, is an important component of the localization
dynamics of the body plan determinant oskar mRNA.
Cells can localize molecules asymmetrically through the combined action of cytoplasmic streaming,
which circulates their fluid contents, and specific anchoring mechanisms.
Streaming also contributes to the distribution of nutrients and organelles such as
chloroplasts in plants, the asymmetric position of the meiotic spindle in mammalian embryos,
and the developmental potential of the zygote, yet little has been known quantitatively about the relationship
between streaming and the motor activity which drives it.
In a collaboration with Dr. Isabel Palacios and her student Dr. Lucy S. Williams (Zoology, Cambridge)
we have used Particle Image Velocimetry (PIV)
to quantify the statistical properties of Kinesin-dependent
streaming during mid-oogenesis in Drosophila.
We find that streaming can be used to detect subtle changes in Kinesin activity and
that the flows reflect the architecture of the microtubule cytoskeleton.
Furthermore, based on characterization of the rheology of the cytoplasm in vivo, we
establish estimates of the number of Kinesins required to drive the observed streaming.
Using this in vivo data as the basis of a model for transport, we suggest that the disordered
character of transport at mid-oogenesis, as revealed by streaming, is an important component of the localization
dynamics of the body plan determinant oskar mRNA.
[back to overview]
Spontaneous Circulation in Confined Active Suspensions
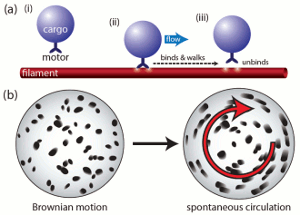 Many active fluid systems encountered in biology are set in total geometric confinement. Cytoplasmic
streaming in plant cells is a prominent and ubiquitous example, in which cargo-carrying molecular
motors move along polymer filaments and generate cell-scale flow. When filaments are not fixed to the
cell periphery, a situation found both in vivo and in vitro, we observe that the basic
dynamics of streaming are closely related to those of a nonmotile stresslet suspension. Under this model,
we have demonstrated that confinement makes possible a
stable circulating state; a linear stability analysis reveals an activity threshold for spontaneous
autocirculation. Numerical analysis of the longtime
behavior reveals a phenomenon akin to defect separation in nematic liquid crystals and a high-activity
bifurcation to an oscillatory regime. These observations provide a possible interpretation to the
experiments of Yotsuyanagi from 1953, in which drops of cytoplasm extracted from the aquatic plant
Chara were observed over time to undergo a transition from disordered random internal motion to organized
persistent circulation.
Many active fluid systems encountered in biology are set in total geometric confinement. Cytoplasmic
streaming in plant cells is a prominent and ubiquitous example, in which cargo-carrying molecular
motors move along polymer filaments and generate cell-scale flow. When filaments are not fixed to the
cell periphery, a situation found both in vivo and in vitro, we observe that the basic
dynamics of streaming are closely related to those of a nonmotile stresslet suspension. Under this model,
we have demonstrated that confinement makes possible a
stable circulating state; a linear stability analysis reveals an activity threshold for spontaneous
autocirculation. Numerical analysis of the longtime
behavior reveals a phenomenon akin to defect separation in nematic liquid crystals and a high-activity
bifurcation to an oscillatory regime. These observations provide a possible interpretation to the
experiments of Yotsuyanagi from 1953, in which drops of cytoplasm extracted from the aquatic plant
Chara were observed over time to undergo a transition from disordered random internal motion to organized
persistent circulation.
[back to overview]
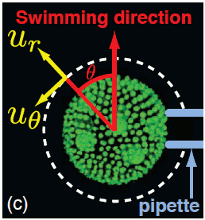 From unicellular ciliates to the respiratory epithelium, carpets of cilia display metachronal waves,
long-wavelength phase modulations of the beating cycles. A large body of theoretical work over the past few
decades has suggested that these waves may arise from
hydrodynamic coupling between the beating flagella. Experimental study of this phenomenon has been limited by a
lack of organisms for which the flagella and the flows they create can be visualized with ease.
Using time-resolved particle image velocimetry and micropipette manipulation, we
report the
discovery of metachronal waves on the surface of the colonial alga Volvox carteri, whose large
size and ease of growth and visualization make it an ideal model organism for these studies. The flagella of
Volvox are relatively far apart compared to the celebrated ciliate Paramecium, and thus more nearly
in the weak-coupling limit amenable to theory. V. carteri robustly
displays symplectic metachronal waves, those for which the wave propagation direction is that of the power stroke of
each flagellum. An elastohydrodynamic model
of weakly coupled compliant oscillators, recast as interacting phase oscillators, reveals that orbit compliance
can produce fast, robust synchronization in a manner essentially independent of boundary conditions, and
offers an intuitive understanding of a possible mechanism leading to the emergence of metachronal waves.
From unicellular ciliates to the respiratory epithelium, carpets of cilia display metachronal waves,
long-wavelength phase modulations of the beating cycles. A large body of theoretical work over the past few
decades has suggested that these waves may arise from
hydrodynamic coupling between the beating flagella. Experimental study of this phenomenon has been limited by a
lack of organisms for which the flagella and the flows they create can be visualized with ease.
Using time-resolved particle image velocimetry and micropipette manipulation, we
report the
discovery of metachronal waves on the surface of the colonial alga Volvox carteri, whose large
size and ease of growth and visualization make it an ideal model organism for these studies. The flagella of
Volvox are relatively far apart compared to the celebrated ciliate Paramecium, and thus more nearly
in the weak-coupling limit amenable to theory. V. carteri robustly
displays symplectic metachronal waves, those for which the wave propagation direction is that of the power stroke of
each flagellum. An elastohydrodynamic model
of weakly coupled compliant oscillators, recast as interacting phase oscillators, reveals that orbit compliance
can produce fast, robust synchronization in a manner essentially independent of boundary conditions, and
offers an intuitive understanding of a possible mechanism leading to the emergence of metachronal waves.
[back to overview]
Antiphase Flagellar Synchronization
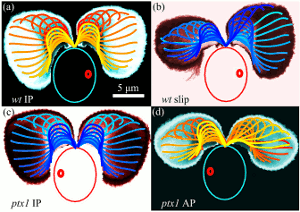 Groups of beating flagella or cilia often synchronize so that
neighboring filaments have identical frequencies and phases. A prime
example is provided by the unicellular biflagellate
Chlamydomonas reinhardtii, which typically displays synchronous
in-phase beating in a low-Reynolds number version of
breaststroke swimming. It is known that steering by flagella during
phototaxis is a consequence of `flagellar dominance', namely that the
two flagella exhibit different responses to intracellular calcium levels
produced by the photoresponse. The mutant ptx1 is a flagellar
dominance mutant in which the asymmetric response is absent. We
report
the discovery that ptx1, can exhibit synchronization in precise
antiphase, as in the freestyle swimming stroke.
High-speed imaging shows that ptx1 flagella switch
stochastically between in-phase and
antiphase states, and that the latter has a distinct waveform and
significantly higher frequency, both of which are strikingly similar to those found during phase slips that stochastically interrupt in-phase
beating of the wild type. Possible mechanisms underlying these observations are discussed.
Groups of beating flagella or cilia often synchronize so that
neighboring filaments have identical frequencies and phases. A prime
example is provided by the unicellular biflagellate
Chlamydomonas reinhardtii, which typically displays synchronous
in-phase beating in a low-Reynolds number version of
breaststroke swimming. It is known that steering by flagella during
phototaxis is a consequence of `flagellar dominance', namely that the
two flagella exhibit different responses to intracellular calcium levels
produced by the photoresponse. The mutant ptx1 is a flagellar
dominance mutant in which the asymmetric response is absent. We
report
the discovery that ptx1, can exhibit synchronization in precise
antiphase, as in the freestyle swimming stroke.
High-speed imaging shows that ptx1 flagella switch
stochastically between in-phase and
antiphase states, and that the latter has a distinct waveform and
significantly higher frequency, both of which are strikingly similar to those found during phase slips that stochastically interrupt in-phase
beating of the wild type. Possible mechanisms underlying these observations are discussed.
[back to overview]
The Many `Phases' of Coupled Flagella
 In a multitude of life's processes, cilia and flagella are found
indispensable. Recently, the biflagellated chlorophyte alga
Chlamydomonas has become a model organism for the study of
ciliary motility and synchronization.
Here,
we use high-speed, high-resolution imaging of single pipette-held
cells to quantify the rich dynamics exhibited by their flagella.
Underlying this variability in behaviour are biological dissimilarities
between the two flagella -- termed cis and trans, with
respect to a unique eyespot.
With emphasis on the wildtype, we derive limit cycles and phase
parameterizations for self-sustained flagellar oscillations from digitally-tracked flagellar waveforms.
Characterizing interflagellar phase-synchrony via a simple model of
coupled oscillators with noise, we find that during the canonical
swimming breaststroke the cis flagellum is consistently
phase-lagged relative to, whilst remaining robustly
phase-locked with, the trans flagellum.
Transient loss of synchrony, or phase-slippage, may be triggered
stochastically, in which the trans flagellum transitions to a
second mode of beating with attenuated beat-envelope and increased
frequency. Further, exploiting this alga's ability for flagellar
regeneration, we mechanically induced removal of one or the other
flagellum of the same cell to reveal a striking disparity between the
beating of the cis vs trans flagellum, in
isolation. These results are evaluated in the context of the dynamic
coordination of Chlamydomonas flagella.
In a multitude of life's processes, cilia and flagella are found
indispensable. Recently, the biflagellated chlorophyte alga
Chlamydomonas has become a model organism for the study of
ciliary motility and synchronization.
Here,
we use high-speed, high-resolution imaging of single pipette-held
cells to quantify the rich dynamics exhibited by their flagella.
Underlying this variability in behaviour are biological dissimilarities
between the two flagella -- termed cis and trans, with
respect to a unique eyespot.
With emphasis on the wildtype, we derive limit cycles and phase
parameterizations for self-sustained flagellar oscillations from digitally-tracked flagellar waveforms.
Characterizing interflagellar phase-synchrony via a simple model of
coupled oscillators with noise, we find that during the canonical
swimming breaststroke the cis flagellum is consistently
phase-lagged relative to, whilst remaining robustly
phase-locked with, the trans flagellum.
Transient loss of synchrony, or phase-slippage, may be triggered
stochastically, in which the trans flagellum transitions to a
second mode of beating with attenuated beat-envelope and increased
frequency. Further, exploiting this alga's ability for flagellar
regeneration, we mechanically induced removal of one or the other
flagellum of the same cell to reveal a striking disparity between the
beating of the cis vs trans flagellum, in
isolation. These results are evaluated in the context of the dynamic
coordination of Chlamydomonas flagella.
[back to overview]
Rheotaxis Facilitates Upstream Navigation of Mammalian Sperm Cells
 A major puzzle in biology is how mammalian sperm determine and maintain
the correct swimming direction during the various phases of the sexual
reproduction process. Whilst chemotaxis is assumed to dominate in the
immediate vicinity of the ovum, it is unclear which biochemical or
physical cues guide spermatozoa on their long journey towards the egg
cell. Currently debated mechanisms range from peristaltic pumping to
temperature sensing (thermotaxis) and direct response to fluid flow
variations (rheotaxis), but little is known quantitatively about their
relative importance. We report the first quantitative experimental
study
of mammalian sperm rheotaxis. Using microfluidic devices, we
investigate systematically the swimming behavior of human and bull
sperm over the whole range of physiologically relevant shear rates
and viscosities. Our measurements show that the interplay of fluid shear,
steric surface-interactions and chirality of the flagellar beat leads
to a stable upstream spiraling motion of sperm cells, thus providing a
generic and robust rectification mechanism to support mammalian
fertilisation. To rationalise these findings, we identify a minimal
mathematical model that is capable of describing quantitatively the
experimental observations. The combined experimental and theoretical
evidence supports the hypothesis that the shape and beat patterns of
mammalian sperm cells have evolved to optimally exploit rheotaxis for
long-distance navigation.
A major puzzle in biology is how mammalian sperm determine and maintain
the correct swimming direction during the various phases of the sexual
reproduction process. Whilst chemotaxis is assumed to dominate in the
immediate vicinity of the ovum, it is unclear which biochemical or
physical cues guide spermatozoa on their long journey towards the egg
cell. Currently debated mechanisms range from peristaltic pumping to
temperature sensing (thermotaxis) and direct response to fluid flow
variations (rheotaxis), but little is known quantitatively about their
relative importance. We report the first quantitative experimental
study
of mammalian sperm rheotaxis. Using microfluidic devices, we
investigate systematically the swimming behavior of human and bull
sperm over the whole range of physiologically relevant shear rates
and viscosities. Our measurements show that the interplay of fluid shear,
steric surface-interactions and chirality of the flagellar beat leads
to a stable upstream spiraling motion of sperm cells, thus providing a
generic and robust rectification mechanism to support mammalian
fertilisation. To rationalise these findings, we identify a minimal
mathematical model that is capable of describing quantitatively the
experimental observations. The combined experimental and theoretical
evidence supports the hypothesis that the shape and beat patterns of
mammalian sperm cells have evolved to optimally exploit rheotaxis for
long-distance navigation.
[back to overview]
Fluid Flows Created by Swimming Bacteria Drive Self-Organization in Confined Suspensions
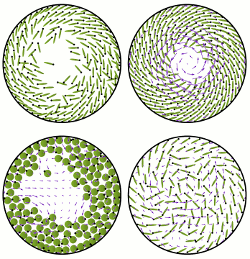 Concentrated suspensions of swimming microorganisms and other forms of
active matter are known to display complex, self-organized spatio-temporal
patterns on scales large compared to those of the individual motile units.
Despite intensive experimental and theoretical study, it has remained
unclear the extent to which the hydrodynamic flows generated by
swimming cells, rather than purely steric interactions between them,
drive the self-organization. We utilize the recent discovery of
a spiral-vortex state in confined suspensions of
B. subtilis to
study this issue in detail.
Those experiments showed that if the radius of confinement in a
thin cylindrical chamber is below a critical value the suspension will
spontaneously form a steady single-vortex state encircled by a counter-rotating cell boundary layer, with
spiral cell orientation within the vortex. Left unclear, however, was the flagellar orientation, and hence the
cell swimming direction, within the spiral vortex.
Here, using a fast simulation method that captures oriented cell-cell
and cell-fluid interactions in a minimal model of discrete-particle systems, we predict the striking, counterintuitive result
that in the presence of
collectively-generated fluid motion the cells within the spiral vortex actually swim upstream against those flows.
This is then confirmed by new experiments reported here, which include measurements of flagella bundle orientation and
cell tracking in the self-organized state. These results highlight the complex interplay between cell orientation and
hydrodynamic flows in concentrated suspensions of microorganisms.
Concentrated suspensions of swimming microorganisms and other forms of
active matter are known to display complex, self-organized spatio-temporal
patterns on scales large compared to those of the individual motile units.
Despite intensive experimental and theoretical study, it has remained
unclear the extent to which the hydrodynamic flows generated by
swimming cells, rather than purely steric interactions between them,
drive the self-organization. We utilize the recent discovery of
a spiral-vortex state in confined suspensions of
B. subtilis to
study this issue in detail.
Those experiments showed that if the radius of confinement in a
thin cylindrical chamber is below a critical value the suspension will
spontaneously form a steady single-vortex state encircled by a counter-rotating cell boundary layer, with
spiral cell orientation within the vortex. Left unclear, however, was the flagellar orientation, and hence the
cell swimming direction, within the spiral vortex.
Here, using a fast simulation method that captures oriented cell-cell
and cell-fluid interactions in a minimal model of discrete-particle systems, we predict the striking, counterintuitive result
that in the presence of
collectively-generated fluid motion the cells within the spiral vortex actually swim upstream against those flows.
This is then confirmed by new experiments reported here, which include measurements of flagella bundle orientation and
cell tracking in the self-organized state. These results highlight the complex interplay between cell orientation and
hydrodynamic flows in concentrated suspensions of microorganisms.
[back to overview]
Flagellar Synchronization Through Direct Hydrodynamic Interactions
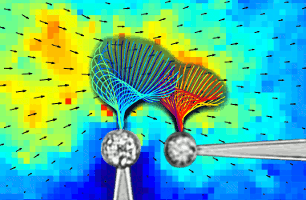 Flows generated by ensembles of flagella are crucial to development,
motility and sensing, but the mechanisms behind this striking coordination
remain unclear. We present novel
experiments in which the two
micropipette-held somatic cells of Volvox carteri, with distinct
intrinsic beating frequencies, are studied by high-speed imaging as a
function of their separation and orientation. Analysis of time series
shows that the interflagellar coupling, constrained by lack of connections
between cells to be hydrodynamical, exhibits a spatial dependence
consistent with theory. At close spacings it produces robust synchrony
for thousands of beats, while at increasing separations synchrony
is degraded by stochastic processes. Manipulation of the relative flagellar
orientation reveals in-phase and antiphase states, consistent with
dynamical theories. Flagellar tracking with exquisite precision
reveals waveform changes that result from hydrodynamic coupling. This
study proves unequivocally that flagella coupled solely through a
fluid can achieve robust synchrony despite differences in their
intrinsic properties.
Flows generated by ensembles of flagella are crucial to development,
motility and sensing, but the mechanisms behind this striking coordination
remain unclear. We present novel
experiments in which the two
micropipette-held somatic cells of Volvox carteri, with distinct
intrinsic beating frequencies, are studied by high-speed imaging as a
function of their separation and orientation. Analysis of time series
shows that the interflagellar coupling, constrained by lack of connections
between cells to be hydrodynamical, exhibits a spatial dependence
consistent with theory. At close spacings it produces robust synchrony
for thousands of beats, while at increasing separations synchrony
is degraded by stochastic processes. Manipulation of the relative flagellar
orientation reveals in-phase and antiphase states, consistent with
dynamical theories. Flagellar tracking with exquisite precision
reveals waveform changes that result from hydrodynamic coupling. This
study proves unequivocally that flagella coupled solely through a
fluid can achieve robust synchrony despite differences in their
intrinsic properties.
[back to overview]
How a Volvox Embryo Turns Itself Inside Out
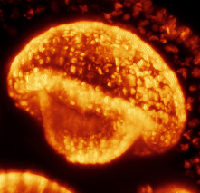 Deformations of cell sheets are ubiquitous in early animal development, often arising from a complex
and poorly understood interplay of cell shape changes, division, and migration.
A prime example of this type of phenomenon is gastrulation, the process in early embryonic
development in which an initially spherical mass of cells develops an invagination that leads
eventually to the formation of the gastric system, and a change of topology to toroidal.
In order to begin to unravel the separate contributions to such folding events, we
have explored perhaps
the simplest example of cell sheet folding: the "inversion" process of the algal genus Volvox, during which
spherical embryos turn themselves inside out through a process hypothesized to arise from cell shape
changes alone. We have used light sheet microscopy to obtain the first four-dimensional (3 space + time)
visualizations of so-called type-B inversion in the species Volvox globator, from which it is possible make detailed quantitative
measurements of many relevant geometry quantities. In this work, we also proposed the first
mathematical theory of this process, in which cell shape changes appear as local
variations of intrinsic curvature, contraction and stretching of a thin elastic shell.
Our results support a scenario
in which these active processes function in a defined spatiotemporal manner to enable inversion.
Deformations of cell sheets are ubiquitous in early animal development, often arising from a complex
and poorly understood interplay of cell shape changes, division, and migration.
A prime example of this type of phenomenon is gastrulation, the process in early embryonic
development in which an initially spherical mass of cells develops an invagination that leads
eventually to the formation of the gastric system, and a change of topology to toroidal.
In order to begin to unravel the separate contributions to such folding events, we
have explored perhaps
the simplest example of cell sheet folding: the "inversion" process of the algal genus Volvox, during which
spherical embryos turn themselves inside out through a process hypothesized to arise from cell shape
changes alone. We have used light sheet microscopy to obtain the first four-dimensional (3 space + time)
visualizations of so-called type-B inversion in the species Volvox globator, from which it is possible make detailed quantitative
measurements of many relevant geometry quantities. In this work, we also proposed the first
mathematical theory of this process, in which cell shape changes appear as local
variations of intrinsic curvature, contraction and stretching of a thin elastic shell.
Our results support a scenario
in which these active processes function in a defined spatiotemporal manner to enable inversion.
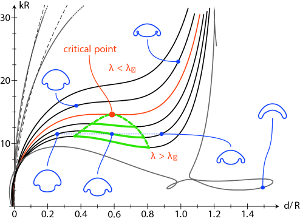 Subsequent work on type B inversion explored in detail the
continuum model, which is based on the biological features of cell shape changes and changes in
the location of intercellular cytoplasmic bridges, which hold the sheet together. When those
bridges connect the midpoints of adjacent cells in a sheet, then cells that become tall and thin
produce contraction of the cell sheet area, while those that become short and squat produce
expansion. When cells become elongated and the bridges migrate to the cell tips, as is known
to be the case during initiation of inversion, then a preferred curvature is created.
We have explored the sequence of quasistatic equilibria that occur when a wave of such changes
progresses around the colony, coupled with anterior expansion and posterior contraction. When
all three occur in the right spatio-temporal sequence, inversion can occur smoothly.
This work also revealed an interesting bifurcation structure in the parameter space, and the possibility
of multiple coexisting shapes. A generalization of the problem to type-A inversion has also been developed,
addressing the bending and peeling of elastic "lips".
Subsequent work on type B inversion explored in detail the
continuum model, which is based on the biological features of cell shape changes and changes in
the location of intercellular cytoplasmic bridges, which hold the sheet together. When those
bridges connect the midpoints of adjacent cells in a sheet, then cells that become tall and thin
produce contraction of the cell sheet area, while those that become short and squat produce
expansion. When cells become elongated and the bridges migrate to the cell tips, as is known
to be the case during initiation of inversion, then a preferred curvature is created.
We have explored the sequence of quasistatic equilibria that occur when a wave of such changes
progresses around the colony, coupled with anterior expansion and posterior contraction. When
all three occur in the right spatio-temporal sequence, inversion can occur smoothly.
This work also revealed an interesting bifurcation structure in the parameter space, and the possibility
of multiple coexisting shapes. A generalization of the problem to type-A inversion has also been developed,
addressing the bending and peeling of elastic "lips".
[back to overview]
Cortical Microtubule Nucleation Can Organise the
Cytoskeleton of Drosophila Oocytes to Define the Anteroposterior Axis
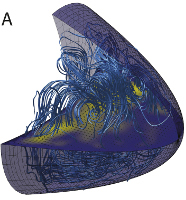 Many cells contain non-centrosomal arrays of microtubules (MTs), but the assembly,
organisation and function of these arrays are poorly understood. We present the first theoretical
model for the non-centrosomal MT cytoskeleton in Drosophila oocytes, in which bicoid and oskar
mRNAs become localised to establish the anterior-posterior body axis. Constrained by experimental
measurements, the model shows that a simple gradient of cortical MT nucleation is sufficient to
reproduce the observed MT distribution, cytoplasmic flow patterns and localisation of oskar and
naive bicoid mRNAs. Our simulations exclude a major role for cytoplasmic flows in localisation and
reveal an organisation of the MT cytoskeleton that is more ordered than previously thought.
Furthermore, modulating cortical MT nucleation induces a bifurcation in cytoskeletal organisation
that accounts for the phenotypes of polarity mutants. Thus, our three-dimensional model explains
many features of the MT network and highlights the importance of differential cortical MT nucleation
for axis formation. Many cells contain non-centrosomal arrays of microtubules (MTs), but the assembly,
organisation and function of these arrays are poorly understood. We present the first theoretical
model for the non-centrosomal MT cytoskeleton in Drosophila oocytes, in which bicoid and oskar
mRNAs become localised to establish the anterior-posterior body axis. Constrained by experimental
measurements, the model shows that a simple gradient of cortical MT nucleation is sufficient to
reproduce the observed MT distribution, cytoplasmic flow patterns and localisation of oskar and
naive bicoid mRNAs. Our simulations exclude a major role for cytoplasmic flows in localisation and
reveal an organisation of the MT cytoskeleton that is more ordered than previously thought.
Furthermore, modulating cortical MT nucleation induces a bifurcation in cytoskeletal organisation
that accounts for the phenotypes of polarity mutants. Thus, our three-dimensional model explains
many features of the MT network and highlights the importance of differential cortical MT nucleation
for axis formation.
[back to overview]
Motility of Colonial Choanoflagellates and the Statistics of
Aggregate Random Walkers
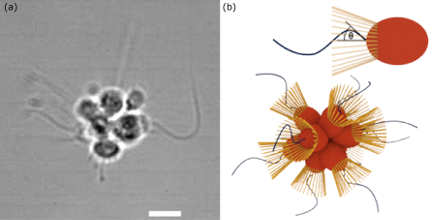 We illuminate the nature of the three-dimensional random walks of microorganisms composed of
individual organisms adhered together. Such aggregate random walkers are typified by choanoflagellates,
eukaryotes that are the closest living relatives of animals and have emerged as as important
model organisms in the study of the evolution of multicellularity. In the colony-forming species Salpingoeca
rosetta, which consists of cells oriented roughly radially, with flagella pointing outwards and the cell
body inward, we show that the beating of each flagellum is stochastic and
uncorrelated with others within the colony. Moreover, the
vectorial sum of the flagellar propulsion forces and torques results in residuals of each that lead to
stochastic helical swimming. A quantitative theory for
these results is presented and species variability discussed.
We illuminate the nature of the three-dimensional random walks of microorganisms composed of
individual organisms adhered together. Such aggregate random walkers are typified by choanoflagellates,
eukaryotes that are the closest living relatives of animals and have emerged as as important
model organisms in the study of the evolution of multicellularity. In the colony-forming species Salpingoeca
rosetta, which consists of cells oriented roughly radially, with flagella pointing outwards and the cell
body inward, we show that the beating of each flagellum is stochastic and
uncorrelated with others within the colony. Moreover, the
vectorial sum of the flagellar propulsion forces and torques results in residuals of each that lead to
stochastic helical swimming. A quantitative theory for
these results is presented and species variability discussed.
[back to overview]
Ferromagnetic and Antiferromagnetic Order in Bacterial Vortex Lattices
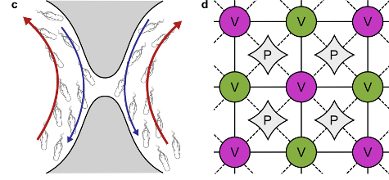 Despite their inherently non-equilibrium nature, living
systems can self-organize in highly ordered collective states
that share striking similarities with the thermodynamic
equilibrium phases of conventional condensed-matter and
fluid systems. Examples range from the liquid-crystal-like
arrangements of bacterial colonies, microbial suspensions
and tissues to the coherent macro-scale dynamics in schools
of fish and flocks of birds. Yet, the generic mathematical
principles that govern the emergence of structure in such
artificial and biological systems are elusive. It is not clear
when, or even whether, well-established theoretical concepts
describing universal thermostatistics of equilibrium systems
can capture and classify ordered states of living matter.
Here,
we connect these two previously disparate regimes: through
microfluidic experiments and mathematical modelling, we
demonstrate that lattices of hydrodynamically coupled
bacterial vortices can spontaneously organize into distinct
patterns characterized by ferro- and antiferromagnetic order.
The coupling between adjacent vortices can be controlled
by tuning the inter-cavity gap widths. The emergence of
opposing order regimes is tightly linked to the existence of
geometry-induced edge currents, reminiscent of those in
quantum systems. Our experimental observations can
be rationalized in terms of a generic lattice field theory,
suggesting that bacterial spin networks belong to the same
universality class as a wide range of equilibrium systems.
Despite their inherently non-equilibrium nature, living
systems can self-organize in highly ordered collective states
that share striking similarities with the thermodynamic
equilibrium phases of conventional condensed-matter and
fluid systems. Examples range from the liquid-crystal-like
arrangements of bacterial colonies, microbial suspensions
and tissues to the coherent macro-scale dynamics in schools
of fish and flocks of birds. Yet, the generic mathematical
principles that govern the emergence of structure in such
artificial and biological systems are elusive. It is not clear
when, or even whether, well-established theoretical concepts
describing universal thermostatistics of equilibrium systems
can capture and classify ordered states of living matter.
Here,
we connect these two previously disparate regimes: through
microfluidic experiments and mathematical modelling, we
demonstrate that lattices of hydrodynamically coupled
bacterial vortices can spontaneously organize into distinct
patterns characterized by ferro- and antiferromagnetic order.
The coupling between adjacent vortices can be controlled
by tuning the inter-cavity gap widths. The emergence of
opposing order regimes is tightly linked to the existence of
geometry-induced edge currents, reminiscent of those in
quantum systems. Our experimental observations can
be rationalized in terms of a generic lattice field theory,
suggesting that bacterial spin networks belong to the same
universality class as a wide range of equilibrium systems.
[back to overview]
Coordinated Beating of Algal Flagella is Mediated by Basal Coupling
 Cilia and flagella often exhibit synchronized behavior; this includes
phase locking, as seen in Chlamydomonas, and metachronal wave
formation in the respiratory cilia of higher organisms. Since the observations by Gray and Rothschild of phase synchrony of nearby
swimming spermatozoa, it has been a working hypothesis that synchrony arises from hydrodynamic interactions between beating filaments. Recent work on the dynamics of physically separated pairs of
flagella isolated from the multicellular alga Volvox has shown that
hydrodynamic coupling alone is sufficient to produce synchrony.
However, the situation is more complex in unicellular organisms bearing few flagella. We show that flagella of Chlamydomonas mutants
deficient in filamentary connections between basal bodies display
markedly different synchronization from the wild type. We perform
micromanipulation on configurations of flagella and conclude that a
mechanism, internal to the cell, must provide an additional flagellar
coupling. In naturally occurring species with 4, 8, or even 16 flagella,
we find diverse symmetries of basal body positioning and of the
flagellar apparatus that are coincident with specific gaits of flagellar
actuation, suggesting that it is a competition between intracellular
coupling and hydrodynamic interactions that ultimately determines
the precise form of flagellar coordination in unicellular algae.
Cilia and flagella often exhibit synchronized behavior; this includes
phase locking, as seen in Chlamydomonas, and metachronal wave
formation in the respiratory cilia of higher organisms. Since the observations by Gray and Rothschild of phase synchrony of nearby
swimming spermatozoa, it has been a working hypothesis that synchrony arises from hydrodynamic interactions between beating filaments. Recent work on the dynamics of physically separated pairs of
flagella isolated from the multicellular alga Volvox has shown that
hydrodynamic coupling alone is sufficient to produce synchrony.
However, the situation is more complex in unicellular organisms bearing few flagella. We show that flagella of Chlamydomonas mutants
deficient in filamentary connections between basal bodies display
markedly different synchronization from the wild type. We perform
micromanipulation on configurations of flagella and conclude that a
mechanism, internal to the cell, must provide an additional flagellar
coupling. In naturally occurring species with 4, 8, or even 16 flagella,
we find diverse symmetries of basal body positioning and of the
flagellar apparatus that are coincident with specific gaits of flagellar
actuation, suggesting that it is a competition between intracellular
coupling and hydrodynamic interactions that ultimately determines
the precise form of flagellar coordination in unicellular algae.
[back to overview]
Directed Collective Motion of Bacteria Under Channel Confinement
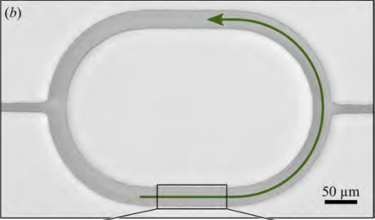 Dense suspensions of swimming bacteria are known to exhibit collective behaviour arising from the
interplay of steric and hydrodynamic interactions. Unconfined suspensions exhibit transient,
recurring vortices and jets, whereas those confined in circular domains may exhibit order in the form
of a spiral vortex. Here, we show that confinement into a long and narrow macroscopic "racetrack"
geometry stabilises bacterial motion to form a steady unidirectional circulation. This motion is
reproduced in simulations of discrete swimmers that reveal the crucial role that bacteria-driven fluid
flows play in the dynamics. In particular, cells close to the channel wall produce strong flows which
advect cells in the bulk against their swimming direction. We examine in detail the transition from a
disordered state to persistent directed motion as a function of the channel width, and show that the
width at the crossover point is comparable to the typical correlation length of swirls seen in the
unbounded system. Our results shed light on the mechanisms driving the collective behaviour of
bacteria and other active matter systems, and stress the importance of the ubiquitous boundaries
found in natural habitats.
Dense suspensions of swimming bacteria are known to exhibit collective behaviour arising from the
interplay of steric and hydrodynamic interactions. Unconfined suspensions exhibit transient,
recurring vortices and jets, whereas those confined in circular domains may exhibit order in the form
of a spiral vortex. Here, we show that confinement into a long and narrow macroscopic "racetrack"
geometry stabilises bacterial motion to form a steady unidirectional circulation. This motion is
reproduced in simulations of discrete swimmers that reveal the crucial role that bacteria-driven fluid
flows play in the dynamics. In particular, cells close to the channel wall produce strong flows which
advect cells in the bulk against their swimming direction. We examine in detail the transition from a
disordered state to persistent directed motion as a function of the channel width, and show that the
width at the crossover point is comparable to the typical correlation length of swirls seen in the
unbounded system. Our results shed light on the mechanisms driving the collective behaviour of
bacteria and other active matter systems, and stress the importance of the ubiquitous boundaries
found in natural habitats.
[back to overview]
Filter-Feeding, Near-field Flows, and the Morphologies of
Colonial Choanoflagellates
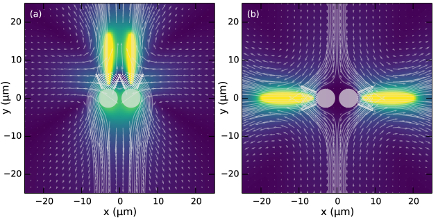 Efficient uptake of prey and nutrients from the environment is an important component in the fitness of
all microorganisms, and its dependence on size may reveal clues to the origins of evolutionary transitions to
multicellularity. Because potential benefits in uptake rates must be viewed in the context of other costs and
benefits of size, such as varying predation rates and the increased metabolic costs associated with larger and more
complex body plans, the uptake rate itself is not necessarily that which is optimized by evolution. Uptake rates
can be strongly dependent on local organism geometry and its swimming speed, providing selective pressure for
particular arrangements. Here, we examine these issues for choanoflagellates, filter-feeding microorganisms that
are the closest relatives of the animals. We explore the different morphological variations of the choanoflagellate
Salpingoeca rosetta, which can exist as a swimming cell, as a sessile thecate cell, and as colonies of cells in
various shapes. In the absence of other requirements and in a homogeneously nutritious environment, we find that
the optimal strategy to maximize filter-feeding by the collar of microvilli is to swim fast, which favors swimming
unicells. In large external flows, the sessile thecate cell becomes advantageous. Effects of prey diffusion are
discussed and also found to be to the advantage of the swimming unicell.
Efficient uptake of prey and nutrients from the environment is an important component in the fitness of
all microorganisms, and its dependence on size may reveal clues to the origins of evolutionary transitions to
multicellularity. Because potential benefits in uptake rates must be viewed in the context of other costs and
benefits of size, such as varying predation rates and the increased metabolic costs associated with larger and more
complex body plans, the uptake rate itself is not necessarily that which is optimized by evolution. Uptake rates
can be strongly dependent on local organism geometry and its swimming speed, providing selective pressure for
particular arrangements. Here, we examine these issues for choanoflagellates, filter-feeding microorganisms that
are the closest relatives of the animals. We explore the different morphological variations of the choanoflagellate
Salpingoeca rosetta, which can exist as a swimming cell, as a sessile thecate cell, and as colonies of cells in
various shapes. In the absence of other requirements and in a homogeneously nutritious environment, we find that
the optimal strategy to maximize filter-feeding by the collar of microvilli is to swim fast, which favors swimming
unicells. In large external flows, the sessile thecate cell becomes advantageous. Effects of prey diffusion are
discussed and also found to be to the advantage of the swimming unicell.
[back to overview]
Elastohydrodynamic Synchronization of Adjacent Beating Flagella
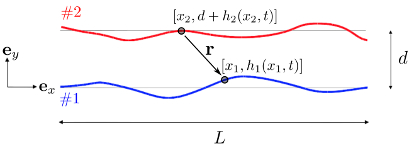 It is now well established that nearby beating pairs of eukaryotic flagella or cilia
typically synchronize in phase. A substantial body of evidence supports the hypothesis
that hydrodynamic coupling between the active filaments, combined with waveform
compliance, provides a robust mechanism for synchrony. This elastohydrodynamic
mechanism has been incorporated into bead-spring models in which the beating flagella
are represented by microspheres tethered by radial springs as they are driven about orbits
by internal forces. While these low-dimensional models reproduce the phenomenon of
synchrony, their parameters are not readily relatable to those of the filaments they represent.
More realistic models, which reflect the underlying elasticity of the axonemes and the active
force generation, take the form of fourth-order nonlinear partial differential equations
(PDEs). While computational studies have shown the occurrence of synchrony, the effects
of hydrodynamic coupling between nearby filaments governed by such continuum models
have been examined theoretically only in the regime of interflagellar distances d large
compared to flagellar length L. Yet in many biological situations d/L≪1. Here we
present an asymptotic analysis of the hydrodynamic coupling between two extended
filaments in the regime d/L≪1 and find that the form of the coupling is independent
of the microscopic details of the internal forces that govern the motion of the individual
filaments. The analysis is analogous to that yielding the localized induction approximation
for vortex filament motion, extended to the case of mutual induction. In order to understand
how the elastohydrodynamic coupling mechanism leads to synchrony of extended objects,
we introduce a heuristic model of flagellar beating. The model takes the form of a single
fourth-order nonlinear PDE whose form is derived from symmetry considerations, the
physics of elasticity, and the overdamped nature of the dynamics. Analytical and numerical
studies of this model illustrate how synchrony between a pair of filaments is achieved
through the asymptotic coupling.
It is now well established that nearby beating pairs of eukaryotic flagella or cilia
typically synchronize in phase. A substantial body of evidence supports the hypothesis
that hydrodynamic coupling between the active filaments, combined with waveform
compliance, provides a robust mechanism for synchrony. This elastohydrodynamic
mechanism has been incorporated into bead-spring models in which the beating flagella
are represented by microspheres tethered by radial springs as they are driven about orbits
by internal forces. While these low-dimensional models reproduce the phenomenon of
synchrony, their parameters are not readily relatable to those of the filaments they represent.
More realistic models, which reflect the underlying elasticity of the axonemes and the active
force generation, take the form of fourth-order nonlinear partial differential equations
(PDEs). While computational studies have shown the occurrence of synchrony, the effects
of hydrodynamic coupling between nearby filaments governed by such continuum models
have been examined theoretically only in the regime of interflagellar distances d large
compared to flagellar length L. Yet in many biological situations d/L≪1. Here we
present an asymptotic analysis of the hydrodynamic coupling between two extended
filaments in the regime d/L≪1 and find that the form of the coupling is independent
of the microscopic details of the internal forces that govern the motion of the individual
filaments. The analysis is analogous to that yielding the localized induction approximation
for vortex filament motion, extended to the case of mutual induction. In order to understand
how the elastohydrodynamic coupling mechanism leads to synchrony of extended objects,
we introduce a heuristic model of flagellar beating. The model takes the form of a single
fourth-order nonlinear PDE whose form is derived from symmetry considerations, the
physics of elasticity, and the overdamped nature of the dynamics. Analytical and numerical
studies of this model illustrate how synchrony between a pair of filaments is achieved
through the asymptotic coupling.
[back to overview]
Aerotaxis in the Closest Relatives of Animals
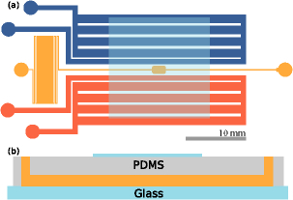 As the closest unicellular relatives of animals, choanoflagellates serve as useful model
organisms for understanding the evolution of animal multicellularity. An important factor in animal
evolution was the increasing ocean oxygen levels in the Precambrian, which are thought to have
influenced the emergence of complex multicellular life. As a first step in addressing these
conditions, we study here the response of the colony-forming choanoflagellate Salpingoeca rosetta
to oxygen gradients. Using a microfluidic device that allows spatio-temporal variations in oxygen
concentrations, we report the discovery that S. rosetta displays positive aerotaxis. Analysis of the
spatial population distributions provides evidence for logarithmic sensing of oxygen, which
enhances sensing in low oxygen neighborhoods. Analysis of search strategy models on the
experimental colony trajectories finds that choanoflagellate aerotaxis is consistent with stochastic
navigation, the statistics of which are captured using an effective continuous version based on
classical run-and-tumble chemotaxis.
As the closest unicellular relatives of animals, choanoflagellates serve as useful model
organisms for understanding the evolution of animal multicellularity. An important factor in animal
evolution was the increasing ocean oxygen levels in the Precambrian, which are thought to have
influenced the emergence of complex multicellular life. As a first step in addressing these
conditions, we study here the response of the colony-forming choanoflagellate Salpingoeca rosetta
to oxygen gradients. Using a microfluidic device that allows spatio-temporal variations in oxygen
concentrations, we report the discovery that S. rosetta displays positive aerotaxis. Analysis of the
spatial population distributions provides evidence for logarithmic sensing of oxygen, which
enhances sensing in low oxygen neighborhoods. Analysis of search strategy models on the
experimental colony trajectories finds that choanoflagellate aerotaxis is consistent with stochastic
navigation, the statistics of which are captured using an effective continuous version based on
classical run-and-tumble chemotaxis.
[back to overview]
Spontaneous Oscillation of Elastic Filaments Induced by Molecular Motors
 It is known from the wave-like motion of microtubules in motility assays that
the piconewton forces that motors produce can be sufficient to bend the filaments. In cellular phenomena such as cytosplasmic streaming, molecular
motors translocate along cytoskeletal filaments, carrying cargo which entrains
fluid. When large numbers of such forced filaments interact through the surrounding fluid, as in particular stages of oocyte development in Drosophila
melanogaster, complex dynamics are observed, but the detailed mechanics
underlying them has remained unclear. Motivated by these observations,
we study perhaps the simplest model for these phenomena: an elastic
filament, pinned at one end, acted on by a molecular motor treated as a
point force. Because the force acts tangential to the filament, no matter what
its shape, this "follower-force" problem is intrinsically non-variational, and
thereby differs fundamentally from Euler buckling, where the force has a fixed
direction, and which, in the low-Reynolds-number regime, ultimately leads to
a stationary, energy-minimizing shape. Through a combination of linear stability
theory, analytical study of a solvable simplified "two-link" model and numerical
studies of the full elastohydrodynamic equations of motion, we elucidate
the Hopf bifurcation that occurs with increasing forcing of a filament, leading
to flapping motion analogous to the high-Reynolds-number oscillations of a
garden hose with a free end.
It is known from the wave-like motion of microtubules in motility assays that
the piconewton forces that motors produce can be sufficient to bend the filaments. In cellular phenomena such as cytosplasmic streaming, molecular
motors translocate along cytoskeletal filaments, carrying cargo which entrains
fluid. When large numbers of such forced filaments interact through the surrounding fluid, as in particular stages of oocyte development in Drosophila
melanogaster, complex dynamics are observed, but the detailed mechanics
underlying them has remained unclear. Motivated by these observations,
we study perhaps the simplest model for these phenomena: an elastic
filament, pinned at one end, acted on by a molecular motor treated as a
point force. Because the force acts tangential to the filament, no matter what
its shape, this "follower-force" problem is intrinsically non-variational, and
thereby differs fundamentally from Euler buckling, where the force has a fixed
direction, and which, in the low-Reynolds-number regime, ultimately leads to
a stationary, energy-minimizing shape. Through a combination of linear stability
theory, analytical study of a solvable simplified "two-link" model and numerical
studies of the full elastohydrodynamic equations of motion, we elucidate
the Hopf bifurcation that occurs with increasing forcing of a filament, leading
to flapping motion analogous to the high-Reynolds-number oscillations of a
garden hose with a free end.
[back to overview]
Mutualism Between Microbial Populations in Structured Environments:
The Role of Geometry in Diffusive Exchanges
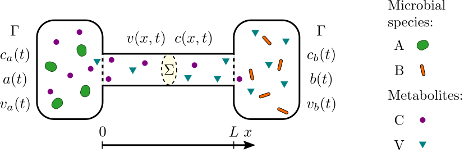 The exchange of diffusive metabolites is known to control the spatial patterns formed by microbial populations,
as revealed by recent studies in the laboratory. However, the matrices used, such as agarose pads, lack the structured
geometry of many natural microbial habitats, including in the soil or on the surfaces of plants or animals. Here we
address the important question of how such geometry may control diffusive exchanges and microbial interaction.
We model mathematically mutualistic interactions within a minimal unit of structure: two growing reservoirs
linked by a diffusive channel through which metabolites are exchanged. The model is applied to study a synthetic
mutualism, experimentally parametrized on a model algal-bacterial co-culture. Analytical and numerical solutions
of the model predict conditions for the successful establishment of remote mutualisms, and how this depends,
often counterintuitively, on diffusion geometry. We connect our findings to understanding complex behavior in
synthetic and naturally occurring microbial communities.
The exchange of diffusive metabolites is known to control the spatial patterns formed by microbial populations,
as revealed by recent studies in the laboratory. However, the matrices used, such as agarose pads, lack the structured
geometry of many natural microbial habitats, including in the soil or on the surfaces of plants or animals. Here we
address the important question of how such geometry may control diffusive exchanges and microbial interaction.
We model mathematically mutualistic interactions within a minimal unit of structure: two growing reservoirs
linked by a diffusive channel through which metabolites are exchanged. The model is applied to study a synthetic
mutualism, experimentally parametrized on a model algal-bacterial co-culture. Analytical and numerical solutions
of the model predict conditions for the successful establishment of remote mutualisms, and how this depends,
often counterintuitively, on diffusion geometry. We connect our findings to understanding complex behavior in
synthetic and naturally occurring microbial communities.
[back to overview]
The Role of Tumbling Frequency and Persistence in Optimal Run-and-Tumble
Chemotaxis
 One of simplest examples of navigation found in nature is run-and-tumble chemotaxis. Tumbles reorient
cells randomly, and cells can drift towards attractants or away from repellents by biasing the frequency of
these events. The post-tumble swimming directions are typically correlated with those prior, as measured
by the variance of the reorientation angle distribution. This variance can range from large, in the case of
bacteria, to so small that tumble events are imperceptible as observed in choanoflagellates. This raises the
question of optimality: why is such a range of persistence observed in nature?
Here, we study persistent
run-and-tumble dynamics, focusing first on the optimization of the linearized chemotactic response
within the 2D parameter space of tumble frequency and angular persistence. Although an optimal
persistence does exist for a given tumble frequency, in the full parameter space there is a continuum of
optimal solutions. Introducing finite tumble times that depend on the persistence can change this picture,
illuminating one possible method for selecting tumble persistence based on species-specific reorientation
dynamics. Moving beyond linear theory we find that optimal chemotactic strengths exist, and that these
maximize reaction when swimming in a wrong direction but have little or no reaction when swimming
with even the slightest projection along the chemoattractant gradient.
One of simplest examples of navigation found in nature is run-and-tumble chemotaxis. Tumbles reorient
cells randomly, and cells can drift towards attractants or away from repellents by biasing the frequency of
these events. The post-tumble swimming directions are typically correlated with those prior, as measured
by the variance of the reorientation angle distribution. This variance can range from large, in the case of
bacteria, to so small that tumble events are imperceptible as observed in choanoflagellates. This raises the
question of optimality: why is such a range of persistence observed in nature?
Here, we study persistent
run-and-tumble dynamics, focusing first on the optimization of the linearized chemotactic response
within the 2D parameter space of tumble frequency and angular persistence. Although an optimal
persistence does exist for a given tumble frequency, in the full parameter space there is a continuum of
optimal solutions. Introducing finite tumble times that depend on the persistence can change this picture,
illuminating one possible method for selecting tumble persistence based on species-specific reorientation
dynamics. Moving beyond linear theory we find that optimal chemotactic strengths exist, and that these
maximize reaction when swimming in a wrong direction but have little or no reaction when swimming
with even the slightest projection along the chemoattractant gradient.
[back to overview]
The Noisy Basis of Morphogenesis: Mechanisms and Mechanics of
Cell Sheet Folding Inferred from Developmental Variability
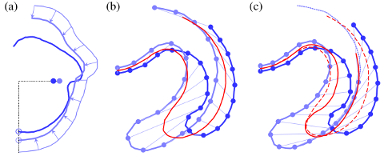 Variability is emerging as an integral part of development. It is therefore imperative to ask
how to access the information contained in this variability. Yet most studies of development
average their observations and, discarding the variability, seek to derive models, biological
or physical, that explain these average observations. Here, we analyse this variability in a
study of cell sheet folding in the green alga Volvox, whose spherical embryos turn themselves inside out in a process sharing invagination, expansion, involution, and peeling of a
cell sheet with animal models of morphogenesis. We generalise our earlier, qualitative
model of the initial stages of inversion by combining ideas from morphoelasticity and shell
theory. Together with three-dimensional visualisations of inversion using light sheet microscopy, this yields a detailed, quantitative model of the entire inversion process. With this
model, we show how the variability of inversion reveals that two separate, temporally uncoupled processes drive the initial invagination and subsequent expansion of the cell sheet.
This implies a prototypical transition towards higher developmental complexity in the volvocine algae and provides proof of principle of analysing morphogenesis based on its
variability.
Variability is emerging as an integral part of development. It is therefore imperative to ask
how to access the information contained in this variability. Yet most studies of development
average their observations and, discarding the variability, seek to derive models, biological
or physical, that explain these average observations. Here, we analyse this variability in a
study of cell sheet folding in the green alga Volvox, whose spherical embryos turn themselves inside out in a process sharing invagination, expansion, involution, and peeling of a
cell sheet with animal models of morphogenesis. We generalise our earlier, qualitative
model of the initial stages of inversion by combining ideas from morphoelasticity and shell
theory. Together with three-dimensional visualisations of inversion using light sheet microscopy, this yields a detailed, quantitative model of the entire inversion process. With this
model, we show how the variability of inversion reveals that two separate, temporally uncoupled processes drive the initial invagination and subsequent expansion of the cell sheet.
This implies a prototypical transition towards higher developmental complexity in the volvocine algae and provides proof of principle of analysing morphogenesis based on its
variability.
[back to overview]
Time-Irreversibility and Criticality in the Motility of a Flagellate
Microorganism
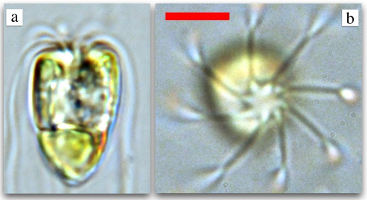 Active living organisms exhibit behavioral variability, partitioning between fast and slow dynamics.
Such variability may be key to generating rapid responses in a heterogeneous, unpredictable environment
wherein cellular activity effects continual exchanges of energy fluxes. We
demonstrate a novel, noninvasive
strategy for revealing nonequilibrium control of swimming - specifically, in an octoflagellate microalga.
These organisms exhibit surprising features of flagellar excitability and mechanosensitivity, which
characterize a novel, time-irreversible "run-stop-shock" motility comprising forward runs, knee-jerk
shocks with dramatic beat reversal, and long stops during which cells are quiescent yet continue to exhibit
submicron flagellar vibrations. Entropy production, associated with flux cycles arising in a reaction graph
representation of the gait-switching dynamics, provides a direct measure of detailed balance violation
in this primitive alga.
Active living organisms exhibit behavioral variability, partitioning between fast and slow dynamics.
Such variability may be key to generating rapid responses in a heterogeneous, unpredictable environment
wherein cellular activity effects continual exchanges of energy fluxes. We
demonstrate a novel, noninvasive
strategy for revealing nonequilibrium control of swimming - specifically, in an octoflagellate microalga.
These organisms exhibit surprising features of flagellar excitability and mechanosensitivity, which
characterize a novel, time-irreversible "run-stop-shock" motility comprising forward runs, knee-jerk
shocks with dramatic beat reversal, and long stops during which cells are quiescent yet continue to exhibit
submicron flagellar vibrations. Entropy production, associated with flux cycles arising in a reaction graph
representation of the gait-switching dynamics, provides a direct measure of detailed balance violation
in this primitive alga.
[back to overview]
Evaporation-Driven Convective Flows in Suspensions of Non-Motile Bacteria
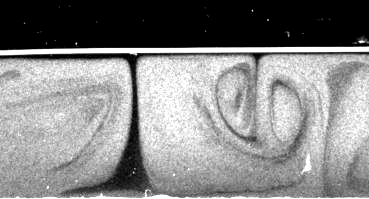 We report a novel form of convection in suspensions of the bioluminescent marine bacterium Photobacterium phosphoreum. Suspensions of these bacteria placed in a chamber
open to the air create persistent luminescent plumes most easily visible when observed
in the dark. These flows are strikingly similar to the classical bioconvection pattern
of aerotactic swimming bacteria, which create an unstable stratification by swimming
upwards to an air-water interface, but they are a puzzle since the strain of P. phosphoreum
used does not express flagella and therefore cannot swim. When microspheres were used
instead of bacteria, similar flow patterns were observed, suggesting that the convective
motion was not driven by bacteria but instead by the accumulation of salt at the airwater interface due to evaporation of the culture medium. Even at room temperature and
humidity, and physiologically relevant salt concentrations, the water evaporation was found
to be sufficient to drive convection patterns. To prove this hypothesis, experiments were
complemented with a mathematical model that aimed to understand the mechanism of
plume formation and the role of salt in triggering the instability. The simplified system
of evaporating salty water was first studied using linear stability analysis, and then with
finite element simulations. A comparison between these three approaches is presented.
While evaporation-driven convection has not been discussed extensively in the context of
biological systems, these results suggest that the phenomenon may be broadly relevant,
particularly in those systems involving microorganisms of limited motility.
We report a novel form of convection in suspensions of the bioluminescent marine bacterium Photobacterium phosphoreum. Suspensions of these bacteria placed in a chamber
open to the air create persistent luminescent plumes most easily visible when observed
in the dark. These flows are strikingly similar to the classical bioconvection pattern
of aerotactic swimming bacteria, which create an unstable stratification by swimming
upwards to an air-water interface, but they are a puzzle since the strain of P. phosphoreum
used does not express flagella and therefore cannot swim. When microspheres were used
instead of bacteria, similar flow patterns were observed, suggesting that the convective
motion was not driven by bacteria but instead by the accumulation of salt at the airwater interface due to evaporation of the culture medium. Even at room temperature and
humidity, and physiologically relevant salt concentrations, the water evaporation was found
to be sufficient to drive convection patterns. To prove this hypothesis, experiments were
complemented with a mathematical model that aimed to understand the mechanism of
plume formation and the role of salt in triggering the instability. The simplified system
of evaporating salty water was first studied using linear stability analysis, and then with
finite element simulations. A comparison between these three approaches is presented.
While evaporation-driven convection has not been discussed extensively in the context of
biological systems, these results suggest that the phenomenon may be broadly relevant,
particularly in those systems involving microorganisms of limited motility.
[back to overview]
II. Natural Pattern Formation
We have a longstanding interest in instabilities and pattern formation in geophysical systems,
dynamics of interfaces, and aspects of the statistical physics of fibrous materials. This work often
combines aspects of differential geometry and nonlinear dynamics.
The Shape of a Ponytail and the Statistical Physics of Hair Fiber Bundles
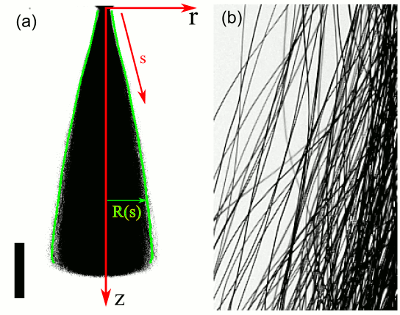 A bundle of hair, whether a paintbrush or a ponytail, adopts a shape determined by the interplay
of the stiffness and weight of the individual fibers and their intrinsic waviness or
curliness. Since a typical bundle may have ten thousand individual hairs, each with a distinct
profile of intrinsic curvatures, the determination of its shape is a problem in the
statistical physics of disordered systems. In work with Patrick B. Warren and Robin C. Ball
we have developed a variational
theory
for the properties of hair bundles. The theory
is based on the local density and mean orientation of hairs, with the effects of random
curvatures subsumed into a local energy functional. This decomposition reveals how the concept of
an `equation of state' of hair appears naturally in the force balance that governs the
equilibrium shape of the bundle. Specializing to long, narrow, axisymmetric bundles,
an ansatz of self-similarity allows the many-body problem to be reduced to an equivalent
single-fiber one for the envelope of the bundle. The interplay between filament stiffness and
weight, and the pressure arising from random curvatures defines two main regimes of ponytail shapes.
This reduction in turn allows the equation of
state of hair to be determined from measurements of ponytail shapes. A robust image-processing
method for determining the three-dimensional shapes of individual hairs and a procedure for
extracting the envelope of a bundle are described in detail. Measurements of those properties
for commercially-available hair switches are presented, and it is shown that the equation of
state thus determined is essentially Hookean, with a spring constant determined by the
bending elasticity and the spectrum of random curvatures of individual fibers.
A bundle of hair, whether a paintbrush or a ponytail, adopts a shape determined by the interplay
of the stiffness and weight of the individual fibers and their intrinsic waviness or
curliness. Since a typical bundle may have ten thousand individual hairs, each with a distinct
profile of intrinsic curvatures, the determination of its shape is a problem in the
statistical physics of disordered systems. In work with Patrick B. Warren and Robin C. Ball
we have developed a variational
theory
for the properties of hair bundles. The theory
is based on the local density and mean orientation of hairs, with the effects of random
curvatures subsumed into a local energy functional. This decomposition reveals how the concept of
an `equation of state' of hair appears naturally in the force balance that governs the
equilibrium shape of the bundle. Specializing to long, narrow, axisymmetric bundles,
an ansatz of self-similarity allows the many-body problem to be reduced to an equivalent
single-fiber one for the envelope of the bundle. The interplay between filament stiffness and
weight, and the pressure arising from random curvatures defines two main regimes of ponytail shapes.
This reduction in turn allows the equation of
state of hair to be determined from measurements of ponytail shapes. A robust image-processing
method for determining the three-dimensional shapes of individual hairs and a procedure for
extracting the envelope of a bundle are described in detail. Measurements of those properties
for commercially-available hair switches are presented, and it is shown that the equation of
state thus determined is essentially Hookean, with a spring constant determined by the
bending elasticity and the spectrum of random curvatures of individual fibers.
[back to overview]
A Ratchet Trap for Leidenfrost Droplets
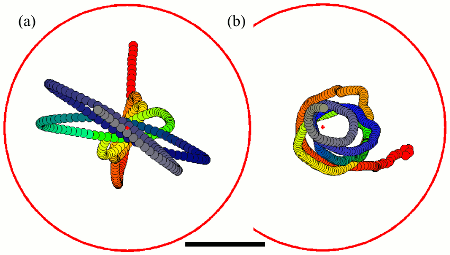 When a Leidenfrost drop is levitated above a surface with parallel asymmetric sawtooth-shaped
ridges it is known to be propelled in a unique direction by the interaction of the
vapour layer with the surface. We have exploited this effect to construct a
`ratchet trap'
for Leidenfrost drops: a surface with concentric circular ridges, each asymmetric in cross
section. A combination of experiment and theory is used to study the dynamics of drops
in these traps, whose centre is a stable fixed point. Numerical analysis of the vapour flow
over a ratchet surface suggests new insights into the mechanism of motion rectification
that are incorporated into the simplest equations of motion for ratchet-driven motion of
a Leidenfrost body; these resemble a central force problem in celestial mechanics with
mass loss and drag. A phase plane analysis of experimental trajectories is used to extract
more detailed information about the ratcheting phenomenon. Orbiting drops are found
to exhibit substantial deformations; those with large internal angular momentum can
even undergo binary fission. Such ratchet traps may thus prove useful in the controlled
study of many properties of Leidenfrost drops.
When a Leidenfrost drop is levitated above a surface with parallel asymmetric sawtooth-shaped
ridges it is known to be propelled in a unique direction by the interaction of the
vapour layer with the surface. We have exploited this effect to construct a
`ratchet trap'
for Leidenfrost drops: a surface with concentric circular ridges, each asymmetric in cross
section. A combination of experiment and theory is used to study the dynamics of drops
in these traps, whose centre is a stable fixed point. Numerical analysis of the vapour flow
over a ratchet surface suggests new insights into the mechanism of motion rectification
that are incorporated into the simplest equations of motion for ratchet-driven motion of
a Leidenfrost body; these resemble a central force problem in celestial mechanics with
mass loss and drag. A phase plane analysis of experimental trajectories is used to extract
more detailed information about the ratcheting phenomenon. Orbiting drops are found
to exhibit substantial deformations; those with large internal angular momentum can
even undergo binary fission. Such ratchet traps may thus prove useful in the controlled
study of many properties of Leidenfrost drops.
[back to overview]
Topological Transitions of Minimal Surfaces
 It is well known that a soap film spanning a looped wire can have
the topology of a Möbius strip and that deformations of the wire
can induce a transformation to a two-sided film, but the process
by which this transformation is achieved has remained unknown.
We have
studied this problem by experiment and theory
and found that the process consists
of a collapse of the film toward the boundary that produces
a previously unrecognized finite-time twist singularity that
changes the linking number of the film's Plateau border and the
centerline of the wire. A movie of this fascinating phenomenon is
here.
We conjecture that it is a general feature
of this type of transition that the singularity always occurs at the
surface boundary. The change in linking number is shown to be a
consequence of a viscous reconnection of the Plateau border at
the moment of the singularity. High-speed imaging of the collapse
dynamics of the film's throat, similar to that of the central opening
of a catenoid, reveals a crossover between two power laws. Far
from the singularity, it is suggested that the collapse is controlled
by dissipation within the fluid film surrounding the wire, whereas
closer to the transition the power law has the classical form arising
from a balance between air inertia and surface tension. Analytical
and numerical studies of minimal surfaces and ruled surfaces are
used to gain insight into the energetics underlying the transition
and the twisted geometry in the neighborhood of the singularity.
It is well known that a soap film spanning a looped wire can have
the topology of a Möbius strip and that deformations of the wire
can induce a transformation to a two-sided film, but the process
by which this transformation is achieved has remained unknown.
We have
studied this problem by experiment and theory
and found that the process consists
of a collapse of the film toward the boundary that produces
a previously unrecognized finite-time twist singularity that
changes the linking number of the film's Plateau border and the
centerline of the wire. A movie of this fascinating phenomenon is
here.
We conjecture that it is a general feature
of this type of transition that the singularity always occurs at the
surface boundary. The change in linking number is shown to be a
consequence of a viscous reconnection of the Plateau border at
the moment of the singularity. High-speed imaging of the collapse
dynamics of the film's throat, similar to that of the central opening
of a catenoid, reveals a crossover between two power laws. Far
from the singularity, it is suggested that the collapse is controlled
by dissipation within the fluid film surrounding the wire, whereas
closer to the transition the power law has the classical form arising
from a balance between air inertia and surface tension. Analytical
and numerical studies of minimal surfaces and ruled surfaces are
used to gain insight into the energetics underlying the transition
and the twisted geometry in the neighborhood of the singularity.
[back to overview]
Boundary Singularities Produced by the Motion of Soap Films
 Recent work has shown that a Möbius strip soap film rendered unstable by deforming its frame changes topology
to that of a disk through a `neck-pinching' boundary singularity. This behaviour is unlike that of the catenoid,
which transitions to two disks through a bulk singularity. It is not yet understood whether the type of singularity is
generally a consequence of the surface topology, nor how this dependence could arise from an equation of motion for the
surface. To address these questions we
investigated
experimentally, computationally, and theoretically the route to singularities
of soap films with different topologies, including a family of punctured Klein bottles. We show that the location of
singularities (bulk or boundary) may depend on the path of the boundary deformation. In the unstable regime the driving
force for soap-film motion is the mean curvature. Thus, the narrowest part of the neck, associated with the shortest
nontrivial closed geodesic of the surface, has the highest curvature and is the fastest-moving. Just before onset of the instability
there exists on the stable surface a shortest closed geodesic, which is the initial condition for evolution of the neck's geodesics,
all of which have the same topological relationship to the frame. We make the plausible conjectures that if the initial geodesic is
linked to the boundary then the singularity will occur at the boundary, whereas if the two are
unlinked initially then the singularity will occur in the bulk. Numerical study of mean curvature flows and experiments support
these conjectures.
Recent work has shown that a Möbius strip soap film rendered unstable by deforming its frame changes topology
to that of a disk through a `neck-pinching' boundary singularity. This behaviour is unlike that of the catenoid,
which transitions to two disks through a bulk singularity. It is not yet understood whether the type of singularity is
generally a consequence of the surface topology, nor how this dependence could arise from an equation of motion for the
surface. To address these questions we
investigated
experimentally, computationally, and theoretically the route to singularities
of soap films with different topologies, including a family of punctured Klein bottles. We show that the location of
singularities (bulk or boundary) may depend on the path of the boundary deformation. In the unstable regime the driving
force for soap-film motion is the mean curvature. Thus, the narrowest part of the neck, associated with the shortest
nontrivial closed geodesic of the surface, has the highest curvature and is the fastest-moving. Just before onset of the instability
there exists on the stable surface a shortest closed geodesic, which is the initial condition for evolution of the neck's geodesics,
all of which have the same topological relationship to the frame. We make the plausible conjectures that if the initial geodesic is
linked to the boundary then the singularity will occur at the boundary, whereas if the two are
unlinked initially then the singularity will occur in the bulk. Numerical study of mean curvature flows and experiments support
these conjectures.
[back to overview]
Instability of a Gravity Current Within a Soap Film
 One of the simplest geometries in which to study fluid flow between two soap films connected by a Plateau border is provided by
a catenoid with a secondary film at its narrowest point. Dynamic variations in the spacing between the two rings supporting
the catenoid lead to fluid flow between the primary and
secondary films. When the rings are moved apart, while keeping their spacing within the overall stability
regime of the films, after a rapid thickening of the secondary film the excess fluid in it starts to drain into the sloped
primary film through the Plateau border at which they meet. This influx of fluid is accommodated by a local thickening of the
primary film. Experiments described
here
show that after this drainage begins the leading edge of the gravity current
becomes linearly unstable to a finite-wavelength fingering instability. A theoretical model based on lubrication theory is used
to explain the mechanism of this instability. The predicted characteristic wavelength of the instability is shown to be in good
agreement with experimental results. Since the gravity current advances into a film of finite, albeit microscopic, thickness this
situation is one in which the regularization often invoked to address singularities at the nose of a thin film is physically justified.
One of the simplest geometries in which to study fluid flow between two soap films connected by a Plateau border is provided by
a catenoid with a secondary film at its narrowest point. Dynamic variations in the spacing between the two rings supporting
the catenoid lead to fluid flow between the primary and
secondary films. When the rings are moved apart, while keeping their spacing within the overall stability
regime of the films, after a rapid thickening of the secondary film the excess fluid in it starts to drain into the sloped
primary film through the Plateau border at which they meet. This influx of fluid is accommodated by a local thickening of the
primary film. Experiments described
here
show that after this drainage begins the leading edge of the gravity current
becomes linearly unstable to a finite-wavelength fingering instability. A theoretical model based on lubrication theory is used
to explain the mechanism of this instability. The predicted characteristic wavelength of the instability is shown to be in good
agreement with experimental results. Since the gravity current advances into a film of finite, albeit microscopic, thickness this
situation is one in which the regularization often invoked to address singularities at the nose of a thin film is physically justified.
[back to overview]
Coiling, Entrainment, and Synchronization of Viscous Fluid Jets
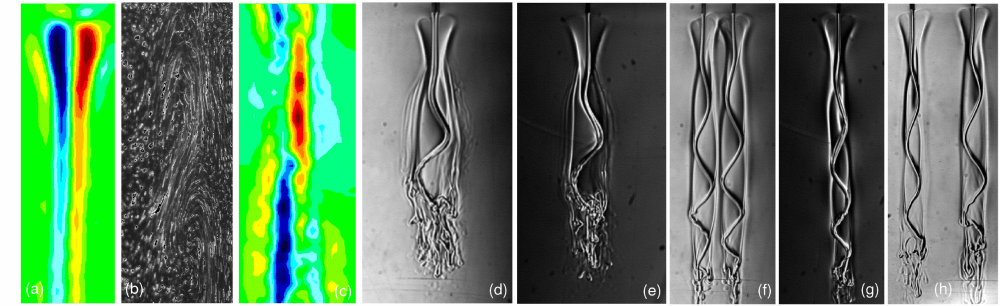 From algal
suspensions to magma upwellings and galactic dynamics one finds fluid
jets which exhibit complex symmetry-breaking instabilities as they are
decelerated by their surroundings. We have
studied
the simplest system that
captures this complexity yet allows direct experimental control; a
saline jet descending through a salinity gradient. The descending
jet coils like a corkscrew within a conduit of viscously entrained
fluid
whose upward recirculation braids the jet. The underlying jet structure
and certain scaling relations can be understood through similarity
solutions to the fluid equations and the physics of Kelvin-Helmholtz
instabilities. The image shows vorticity maps (color) obtained from PIV
with suspended microspheres that illustrate the recirculating flow
within the conduit in a time-averaged manner (left) and instantaneously
(right), and a streak photograph (b) offering another view of the
swirling flows that braid the coiling jet. Panels (d,e) and (f,g)
show from and side views of two interacting jets that are,
respectively,
sharing a single conduit and just further apart than a conduit width.
In the latter case, the jets synchronize as mirror-images.
Panel h reveals that when jets are even further apart they nest
together. More recently we have done extensive numerical and
analytical work on the underlying mechanism of the instability in a
two-dimensional version of this
problem.
From algal
suspensions to magma upwellings and galactic dynamics one finds fluid
jets which exhibit complex symmetry-breaking instabilities as they are
decelerated by their surroundings. We have
studied
the simplest system that
captures this complexity yet allows direct experimental control; a
saline jet descending through a salinity gradient. The descending
jet coils like a corkscrew within a conduit of viscously entrained
fluid
whose upward recirculation braids the jet. The underlying jet structure
and certain scaling relations can be understood through similarity
solutions to the fluid equations and the physics of Kelvin-Helmholtz
instabilities. The image shows vorticity maps (color) obtained from PIV
with suspended microspheres that illustrate the recirculating flow
within the conduit in a time-averaged manner (left) and instantaneously
(right), and a streak photograph (b) offering another view of the
swirling flows that braid the coiling jet. Panels (d,e) and (f,g)
show from and side views of two interacting jets that are,
respectively,
sharing a single conduit and just further apart than a conduit width.
In the latter case, the jets synchronize as mirror-images.
Panel h reveals that when jets are even further apart they nest
together. More recently we have done extensive numerical and
analytical work on the underlying mechanism of the instability in a
two-dimensional version of this
problem.
[back to overview]
Speleothem Morphology
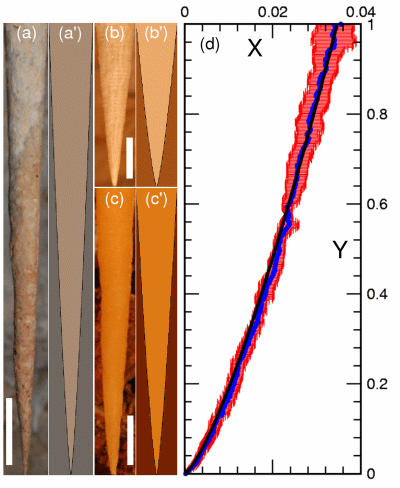 The chemical mechanisms underlying the growth of
cave formations such as stalactites are well-known, yet no theory had
been proposed which successfully accounts for the dynamic evolution
of their shapes. We have developed such a theory
(see also a longer paper)
that considers the interplay of thin-film
fluid
dynamics, calcium carbonate chemistry, and CO2 transport in
the cave to show that stalactites evolve according to a novel local
geometric growth law which exhibits extreme amplification at the tip as
a consequence of the locally-varying fluid layer thickness. Studies of
this model show that a broad class of initial conditions is attracted
to an ideal shape which is strikingly close to a statistical average of
natural stalactites. This ideal shape has the
remarkable property of being completely parameter-free, save for an
overall scaling, as for the Platonic ideals like the circle and the
square. The figure shows (left) three examples of natural
stalactites in Kartchner Caverns (Benson, AZ) and the ideal shape which
most closely matches them. At the right is a composite average
shape (in blue, with statistical uncertainties in red) compared
with our calculated Platonic ideal of stalactites (black). The
agreement is striking. We are currently extending these
ideas to understand non-axisymmetric shapes found in caves and the
surface ripple instability commonly seen.
The chemical mechanisms underlying the growth of
cave formations such as stalactites are well-known, yet no theory had
been proposed which successfully accounts for the dynamic evolution
of their shapes. We have developed such a theory
(see also a longer paper)
that considers the interplay of thin-film
fluid
dynamics, calcium carbonate chemistry, and CO2 transport in
the cave to show that stalactites evolve according to a novel local
geometric growth law which exhibits extreme amplification at the tip as
a consequence of the locally-varying fluid layer thickness. Studies of
this model show that a broad class of initial conditions is attracted
to an ideal shape which is strikingly close to a statistical average of
natural stalactites. This ideal shape has the
remarkable property of being completely parameter-free, save for an
overall scaling, as for the Platonic ideals like the circle and the
square. The figure shows (left) three examples of natural
stalactites in Kartchner Caverns (Benson, AZ) and the ideal shape which
most closely matches them. At the right is a composite average
shape (in blue, with statistical uncertainties in red) compared
with our calculated Platonic ideal of stalactites (black). The
agreement is striking. We are currently extending these
ideas to understand non-axisymmetric shapes found in caves and the
surface ripple instability commonly seen.
[back to overview]
Icicle Morphology
 In light of the results described above on stalactite growth it was natural to wonder how
it is that icicles obtain their characteristic long and pointy shapes, and we have begun to
investigate the growth and melting of icicles as free-boundary problems. Our first
work describes perhaps the simplest synthesis of atmospheric heat
transfer, geometrical considerations, and thin-film fluid dynamics. Crucial to this
analysis is the power-law form of the thermal boundary layer rising upwards adjacent to the
icicle surface. It acts as an insulating thermal blanket whose thickness self-consistently
determines the icicle growth rate. This analysis led to the
discovery of a nonlinear ordinary
differential equation for the shape of a uniformly advancing icicle, the solution to which defines a
parameter-free shape which compares very favorably with that of natural icicles. Away from the tip,
the solution has a power-law form identical to that found for the growth of stalactites by
precipitation of calcium carbonate. This analysis thereby explains why stalactites and icicles are so
similar in form despite the vastly different physics and chemistry of their formation. In addition, a
curious link is noted between the shape so calculated and that found through consideration of only
the thin coating water layer.
In light of the results described above on stalactite growth it was natural to wonder how
it is that icicles obtain their characteristic long and pointy shapes, and we have begun to
investigate the growth and melting of icicles as free-boundary problems. Our first
work describes perhaps the simplest synthesis of atmospheric heat
transfer, geometrical considerations, and thin-film fluid dynamics. Crucial to this
analysis is the power-law form of the thermal boundary layer rising upwards adjacent to the
icicle surface. It acts as an insulating thermal blanket whose thickness self-consistently
determines the icicle growth rate. This analysis led to the
discovery of a nonlinear ordinary
differential equation for the shape of a uniformly advancing icicle, the solution to which defines a
parameter-free shape which compares very favorably with that of natural icicles. Away from the tip,
the solution has a power-law form identical to that found for the growth of stalactites by
precipitation of calcium carbonate. This analysis thereby explains why stalactites and icicles are so
similar in form despite the vastly different physics and chemistry of their formation. In addition, a
curious link is noted between the shape so calculated and that found through consideration of only
the thin coating water layer.
[back to overview]
 More recently, we have studied
in a more controlled fashion the melting of a cylinder of ice in
warm air in order to quantify the
heat-transfer mechanisms controlling the evolution of its shape, which are inherent
in a range of phenomena involving phase change and fluid flow. Motivated by
the initial melting at the top of a flat-topped cylinder of ice, we analyse laminar,
natural convection above a cooled, finite, horizontal plate (or below a heated, finite,
horizontal plate) and show that, to a very good approximation, the partial-differential,
boundary-layer equations can be separated with self-similar vertical profiles scaled by
the boundary-layer thickness. We find that the horizontal evolution of the boundary-layer
thickness is governed by equations describing a steady, viscous gravity current
fed by diffusive entrainment, and therefore describe such flows as diffusive gravity
currents. We first use the predictions of our model to examine previous experimental
results in two dimensions. Our experimental results relating to the melting of ice in
air are then compared with predictions based on our analysis of the axisymmetric
thermal boundary layer. This comparison confirms the vertical thermal structure and
shows that melting is governed in roughly equal measure by heat transfer from the air,
the latent heat of condensation of water vapour, and the net radiative heat transfer
from the surroundings to the ice.
More recently, we have studied
in a more controlled fashion the melting of a cylinder of ice in
warm air in order to quantify the
heat-transfer mechanisms controlling the evolution of its shape, which are inherent
in a range of phenomena involving phase change and fluid flow. Motivated by
the initial melting at the top of a flat-topped cylinder of ice, we analyse laminar,
natural convection above a cooled, finite, horizontal plate (or below a heated, finite,
horizontal plate) and show that, to a very good approximation, the partial-differential,
boundary-layer equations can be separated with self-similar vertical profiles scaled by
the boundary-layer thickness. We find that the horizontal evolution of the boundary-layer
thickness is governed by equations describing a steady, viscous gravity current
fed by diffusive entrainment, and therefore describe such flows as diffusive gravity
currents. We first use the predictions of our model to examine previous experimental
results in two dimensions. Our experimental results relating to the melting of ice in
air are then compared with predictions based on our analysis of the axisymmetric
thermal boundary layer. This comparison confirms the vertical thermal structure and
shows that melting is governed in roughly equal measure by heat transfer from the air,
the latent heat of condensation of water vapour, and the net radiative heat transfer
from the surroundings to the ice.
[back to overview]
Instability of a Möobius Strip Minimal Surface and a Link
with Systolic Geometry
 We describe the first analytically tractable example of an instability of a nonorientable minimal surface
under parametric variation of its boundary. A one-parameter family of incomplete Meeks Möbius surfaces
is defined and shown to exhibit an instability threshold as the bounding curve is opened up from a doublecovering of the circle. Numerical and analytical methods are used to determine the instability threshold
by solution of the Jacobi equation on the double covering of the surface. The unstable eigenmode shows
excellent qualitative agreement with that found experimentally for a closely related surface. A connection is
proposed between systolic geometry and the instability by showing that the shortest noncontractable closed
geodesic on the surface (the systolic curve) passes near the maximum of the unstable eigenmode.
We describe the first analytically tractable example of an instability of a nonorientable minimal surface
under parametric variation of its boundary. A one-parameter family of incomplete Meeks Möbius surfaces
is defined and shown to exhibit an instability threshold as the bounding curve is opened up from a doublecovering of the circle. Numerical and analytical methods are used to determine the instability threshold
by solution of the Jacobi equation on the double covering of the surface. The unstable eigenmode shows
excellent qualitative agreement with that found experimentally for a closely related surface. A connection is
proposed between systolic geometry and the instability by showing that the shortest noncontractable closed
geodesic on the surface (the systolic curve) passes near the maximum of the unstable eigenmode.
[back to overview]
Instabilities and Solitons in Minimal Strips
 We show that highly twisted minimal strips can undergo a nonsingular transition, unlike the singular
transitions seen in the Möbius strip and the catenoid. If the strip is nonorientable, this transition is
topologically frustrated, and the resulting surface contains a helicoidal defect. Through a controlled
analytic approximation, the system can be mapped onto a scalar ?4 theory on a nonorientable line bundle
over the circle, where the defect becomes a topologically protected kink soliton or domain wall, thus
establishing their existence in minimal surfaces. Demonstrations with soap films confirm these results and
show how the position of the defect can be controlled through boundary deformation.
We show that highly twisted minimal strips can undergo a nonsingular transition, unlike the singular
transitions seen in the Möbius strip and the catenoid. If the strip is nonorientable, this transition is
topologically frustrated, and the resulting surface contains a helicoidal defect. Through a controlled
analytic approximation, the system can be mapped onto a scalar ?4 theory on a nonorientable line bundle
over the circle, where the defect becomes a topologically protected kink soliton or domain wall, thus
establishing their existence in minimal surfaces. Demonstrations with soap films confirm these results and
show how the position of the defect can be controlled through boundary deformation.
[back to overview]
Theory of Shape-Shifting Droplets
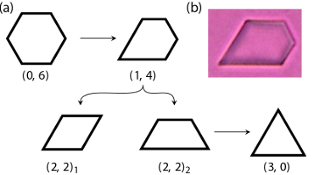 Recent studies of cooled oil emulsion droplets uncovered transformations into a host of flattened shapes
with straight edges and sharp corners, driven by a partial phase transition of the bulk liquid phase. Here, we
explore theoretically the simplest geometric competition between this phase transition and surface tension
in planar polygons and recover the observed sequence of shapes and their statistics in qualitative agreement
with experiments. Extending the model to capture some of the three-dimensional structure of the droplets,
we analyze the evolution of protrusions sprouting from the vertices of the platelets and the topological
transition of a puncturing planar polygon.
Recent studies of cooled oil emulsion droplets uncovered transformations into a host of flattened shapes
with straight edges and sharp corners, driven by a partial phase transition of the bulk liquid phase. Here, we
explore theoretically the simplest geometric competition between this phase transition and surface tension
in planar polygons and recover the observed sequence of shapes and their statistics in qualitative agreement
with experiments. Extending the model to capture some of the three-dimensional structure of the droplets,
we analyze the evolution of protrusions sprouting from the vertices of the platelets and the topological
transition of a puncturing planar polygon.
[back to overview]
Why Clothes Don't Fall Apart: Tension Transmission in Staple Yarns
 The problem of how staple yarns, those composed of myriads of short fibers, transmit tension
was first described by Galileo in the context of ropes in his
Dialogues Concerning Two New Sciences (1638). We examine
the mechanics of this problem using abstract models in which the
Amontons-Coulomb friction laws yield a linear programing (LP) problem for the tensions in the fiber
elements. We find there is a percolation transition such that above the percolation threshold the transmitted
tension is in principle unbounded. We determine that the mean slack in the LP constraints is a suitable order
parameter to characterize this supercritical state. We argue the mechanism is generic, and in practical terms,
it corresponds to a switch from a ductile to a brittle failure mode accompanied by a significant increase in
mechanical strength.
The problem of how staple yarns, those composed of myriads of short fibers, transmit tension
was first described by Galileo in the context of ropes in his
Dialogues Concerning Two New Sciences (1638). We examine
the mechanics of this problem using abstract models in which the
Amontons-Coulomb friction laws yield a linear programing (LP) problem for the tensions in the fiber
elements. We find there is a percolation transition such that above the percolation threshold the transmitted
tension is in principle unbounded. We determine that the mean slack in the LP constraints is a suitable order
parameter to characterize this supercritical state. We argue the mechanism is generic, and in practical terms,
it corresponds to a switch from a ductile to a brittle failure mode accompanied by a significant increase in
mechanical strength.
[back to overview]
|